研究をふりかえりみて
研究の動機、なぜ? 研究のあゆみ 研究の結果と評価について
私の研究の概要は「自己紹介」で書きましたが、ここでは多少詳細に、その動機、これまでの歩み、成果と評価、そして今つづけているの仕事を書きます。
目次
1 研究のあゆみと姿
2 研究の動機、問題意識と自分の立ち位置
3 研究の到達点と評価
4 研究テーマと研究のあゆみ
研究テーマの考え方
5 魚の歯に関して
6 イヌのエナメル質の研究
7 日本大学の松戸歯学部での研究
8 結晶の研究
9 仕事のまとめ
10 反省 残念なこと
1 研究のあゆみと姿
研究の過程は大きく4つの時期に分かれます。
第一の時代は解剖の修業をした東京医科歯科時代と埼玉医科大学時代、前者は解剖学と組織学あるいは細胞学という基本の勉強、後者は発生学の勉強を主にしました。同時に長鼻類の仕事やエナメル質の仕事をすこしずつ進めました。
第二の時期は、日本大学の松戸歯学部の時代で、前半の講師、助教授時代、前半はエナメル質の形成とエナメル質構造の比較をしらべました。
第三の時期、松戸での後半は免疫組織化学などを借りてより深く構造の違いはどこから来るのかを調べました。
第四の時期は、定年の数年前から現在までです。ここではこれまでの成果をまとめ、法則を普遍化し、進化の問題に踏み込んでいます。
概観すれば、師匠の井尻さんへ「進化の研究をしたい」とお願いして入門し、まず解剖学を、と言われてから40年、最初の課題に戻った、ということです。なんというか不思議な因縁を感じます。
忘れられない大学院生とスタッフの力!
振り返りみれば、特に松戸に来てからスタッフの力、とくに定年前20年間は大学院生の力が大きかったと感謝しています。大学院生には厳しくあたりましたが、それでも彼らの創意と底力は凄く、つぎつぎと新しい成果がでました。なぜか失敗が少なく、予想する結果がぞくぞくと生まれました。彼等がいなければ現在の成果までたどり着けなかったと、感謝し反省しています。その意味でみんなの成果と考えています。
研究の原点とも言うべき問題意識の第一は、研究を始めてから今日まで変わらずに抱いていた疑問、問題意識があります。それがわたくしの研究の原点だしこれまで続けてきた原動力だと感じています。
問題意識は、歯の細胞とか組織の研究を進めていくうえで、細胞、組織、歯の構造、歯の形の相互関連性はどうなっているのか、という点です。これはヒエラルキーつまり階層の間の相互の関係です。 解剖の講義は、細胞学、組織学、系統解剖、各器官の発生が別々に、つまり階層別に行われ、この身体を、器官を、組織を構成する細胞はこうなっている、体をこう構成している、と詳しく講義されます。(当時の東京医科歯科大学の解剖の先生たちは超一流、同僚もみな秀才の塊まりだったため、このなかで凡才の私は着いてゆくのが必死のありさまでしたが・・・・・)
しかし、わたくしの目指したエナメル小柱の断面の形の動物種での違いは、何が原因でそうなるのか、歯の組織や歯の形態とどのような関係にあるのか、がどこにも書かれていませんでした。つまり未解決だったのです。これにたいする答えがある程度わかったのは「歯の形態形成原論」をまとめたとき、ヒエラルキーつまり階層性の問題を分析したときです。階層間の調整の問題だったのです。鈍才の私には約40年間かかりました。
第二点は、今現在、進化の分析に当たり自分の仕事や知識が「大海の一滴」だと実感し、これを乗り越え、補うのは哲学の利用しかないと考えるに到ったことです。
哲学についての最初の出会いは、姉の下宿していた吉祥寺の古書店(?)で甘粕石介さんの「ヘーゲル弁証法」だったか?を買い、「正反合」「否定の否定」との記述へ感動して、紙に書き出し机の前に張っておりました。どうして高校生の自分がこの本を購入したのか定かではありませんが、秀才の親友で亡くなった渡辺義廣君に「俺はいつも「一」とかいた紙を持っていて、これが何を意味するか考えている」といわれて衝撃を受けたからかもしれません。
しかし、運命は異なもので、旧姓甘粕の見田石介さんとは師匠の井尻正二先生を介して知ることになります。
私は、他の人からの評価はできるだけ理解して受けるようにしていますが、基本的には自分なりに反省し評価(自己評価)するようにしています。なぜなら学問は多様であり、評価も立ち位置によって多様です。ちょっと分野が違うと理解が困難なことが沢山あります。自分ではあまり評価を下さないようにしています。自分の視点でほかの研究へ評価を下すと評論者のようになってしまうからです。評価は反省材料として必要なのですが、評論者に、師匠の井尻さんが「評論者がなにを産み出すのか!」と一喝されたことをいつも思い出します。
さて私は、研究過程で色々な法則を考え、最終的に進化までたどり着いたのですが、この法則をはじめ私のような進化の分析は他にない、とほんのちょっと自負しています。それは、これらの法則なくして進化を理解できず、ヒトの体を理解できない、と確信を持つからです。ですから不安や疑問はたくさんありながらも、ある程度の達成感を持っています。
これに似た感情を持ったのは、今から四半世紀くらい前でしょうか、師匠の井尻さんから「100種類の動物を見るまで比較解剖とはいえない」と釘を刺され、あの秀才揃いの東京医科歯科大学の解剖学教室で、鈍才の私が右往左往しながら仕事を進め、やっと100種類に近くなり、国内外から問い合わせが来るようになった時です。初めはどうしてこんな問い合わせが来るのかな、と思っていたのですが、100種の比較をした人は他に余りいないとわかり、不安に混じってちょっとした達成感を持ちました。その時以来と言うことになります。
ただし、私の「体制の原理(法則)」や進化の基本法則を理解してくれる人がいるかとい言うと、どうかな?と感じています。たまに海外の友人から「お前の仕事を否定しているのがいるから、論文を見て反論しろ」との連絡を受けますが、このような友人が海外にもいるという感動とともに、ごくごく限られていると感じます。
しかし、今後10年、20年あるいはさらに長期的にみると、このような人がもっと現れると確信しています。なぜなら、上に書いたように私の結論を迂回して、進化や体の理解はできないからです。
私の結論に近いところまで達していたのは、師匠の井尻(正二)さんと三木(成夫)さんです。両師匠ともに鬼籍に入りましたが、私の才能のなさと不甲斐なさによって、生前に議論できなかったことに対し慚愧の念を持っています。(これには後日談があり、議論する人がいなくなってしまったと漏らしたところ、宇宙地質学を提唱されている先輩の小森長生さんから、僕なんか最初からひとりだった、と叱られました。持つのはよき師匠であり、よき先輩です。)
4 研究テーマと研究のあゆみ
まえがき 解剖学教室で・・・
学生時代に「進化」を研究したいと漠然と考えていました。そのころ大森昌衛先生(当時は教育大)の本を読み、糞化石という化石があることを知り、これなら腸の形が分かり今の動物と同様な研究方法で進化を扱えるのではなにかと考えました。
そこで当時、古生物学でも生物学と同様の実験が必要と歯の移植実験などをしていて、「古生物学論」(いまは科学論と改題)を書かれていた井尻正二先生に直接電話して、相談しました。その結果、日本には動物の古生物学を扱うのは教育大の大森昌衛先生か京都大学の亀井節夫先生の大学院が良いという話を給わりました。両方とも地質学教室でそれから、初めての地質学のにわか勉強をしました。この勉強には当時の教育大学の大学院生の方々にお世話になりました。しかし、お恥ずかしながら両方とも不合格。
その後ちょっと経緯を踏んで医科歯科大学の解剖学教室の助手になりました。当初は三木成夫先生につければ、という話だったのですが、いろいろな事情で歯学部の桐野忠大先生、一条尚先生のもとに入門することになりました。(結果的にはこれが幸運の始まりで、古生物学には欠かせない歯の勉強と研究をすることができ、歯の構造と発生、化石の歯の構造に研究対象を広げることがで、今日の基礎ができました。)
研究テーマ
ここで研究テーマを選ぶ様子に触れておきます。
研究テーマは原則的に各自自由に決めるべきです。しかし、これだけ情報が多いとなにをやってよいか五里霧中の状態になってしまいます。よく論文を勉強してテーマを決めろ、と言う方がいますが一般的な経験で初心者に論文を読みこなすのは無理ではないか、という感想を持っています。そこで指導者に相談することになるのですが・・・。次に書くのは私自身の経験で読まれる方の参考になれば、と考えています。
東京医科歯科大学解剖学教室の研究テーマ
当時、教室の大きなテーマは比較解剖であったため、同僚はそれぞれ魚の歯の研究をすすめており、私はイスズミ(オキナメジナ)という魚の歯の組織発生を担当することになりました。
いやはや教室には同じ年頃の秀才ぞろいで、鈍な私はウロウロしながら仕事を進め、かつ人体解剖、組織、細胞、発生、そして歯の形態、歯の組織を学びつつ教育にもあたる羽目になりました。(当時の解剖学教室は、予科練と同じ月月火水木金金で、休む暇はまずありません、優秀な先輩から、メス研ぎ3年HE(染色技術の基礎)7年、と聞かされ、はじめはメス研ぎの毎日でした・・・。教授が朝来るのが早いので早朝6時半ごろ家を出て7時過ぎに授業準備、帰るのは最終の電車という毎日でした。日曜日にも呼び出されて夜中まで仕事しました。(しかし、このころ勉強したことはよく覚えているのに自分自身で驚いています。)
大切な反省点は、手がけた研究はいずれも後で役立っている、ということです。まず人生にはその時点では無駄だと感じても、無駄はない、無駄なようにみえてもじつは血となり肉になっている、ということです。
その後、みんなで動物を広く分担するということになり、わたくしは哺乳類のイヌの歯の細胞分化・組織発生を研究することになりました。ちなみにほかの方々は、サメ、魚、ワニなどでした。(井尻さんは、ワニが面白かったかも、といわれたのですが、材料が入手しやすいイヌで歯の基本的なことを勉強できれば、と考えたのです。そしてその延長にやがて、両生類や爬虫類ネコやブタ等々、そして脊椎動物の先祖に近いといわれる火屋(ホヤ)の棘や歯、甲殻類の殻の形成や発生を比較することになりました。ここでも無駄がなかったなー、という感想を持っています)。
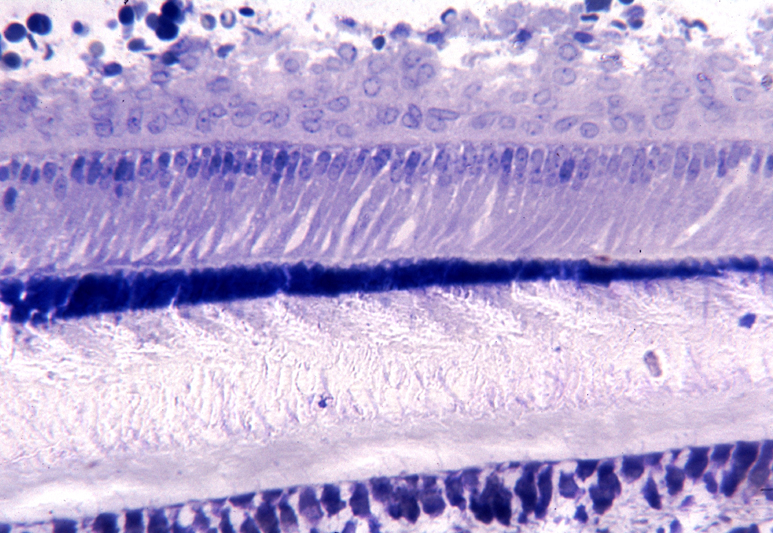
| |
| 濃く青い領域がエナメル質、その上がエナメル芽細胞、下が象牙質一番下の青い点々が象牙芽細胞 |
研究テーマを選ぶのは「好み」でも、研究目的や意義についての理論的バックボーンは必要なことを師匠の井尻さんからおしえていただきました。
研究テーマは、教授に与えられることや、自ら選ぶなど、いろいろありますが、具体的に研究をすすめるのはなかなか難しいものです。勢い先生や先輩に相談することになります。
(余談ですが、当時は、教授のファッショで自分の望む研究をさせてくれない、という話をよく耳にしました。私の経験から、確かにそういう面もあったかもしれませんが、教授の指示で基礎を勉強して、つぎに自分のテーマに向かう、という点が大切だと感じています。)
私は「進化」を研究したいと希望したところ、井尻先生に与えられたテーマは「長鼻類の歯」でした。その理由は「世界的でかつ世界的テーマ」であること、つまり世界に分布していて、世界の人が誰でも知る研究という意味です。(私は世界中の人が知っているためにその価値を世界中の人が理解できる、と理解しています)。
そしてもう一つの意味は、最も進化したものにはその進化過程の要素が含まれているので、進化の「極」から振り返るのがよい、というものでした(後にこれは、マルクスの資本論の研究手法と同じだとわかりましたが・・・)。
当時、化石研究会の中に「象団研」(京都大学の故亀井節夫先生を中心にゾウを研究する目的の研究会)があり、これが改組されて「長鼻類団研」となりました。これに加えていただき先輩の指導の下に研究(特に組織の研究)することになりました。長鼻類というのは、通称であるウゾの仲間全体を指す「長鼻目」という分類単位の名前で、研究団体のなまえもこれに従ったと聞いています。
(反省 この研究グループに参加させていただいた時は忘れられない思い出があります。化石を対象に生化学の研究をしていた(現在はない資源研究所の)故藤原隆代さんという先輩から長鼻類の組織標本を引き継いだときのことです。研究会で井尻さんが「組織の研究をするのはだれがよいか?」と私の返事を期待して話したのに、ボーッとしていて答えなかったのです。すかさず私にむかって藤原さんが、じっと目を見つめて(怖かったですね)「自分から申し出ないような人は期待できない!」と叱られました。はっと気が付いて「私でよければ・・・」とおずおずと申し出て、皆さんは「まあ仕方がないか・・・」というような雰囲気でこの仕事を任されたのです。先輩たちは研究に真剣でかつテーマを深く理解していたのだな、と感謝と自分の不甲斐なさの反省です。)
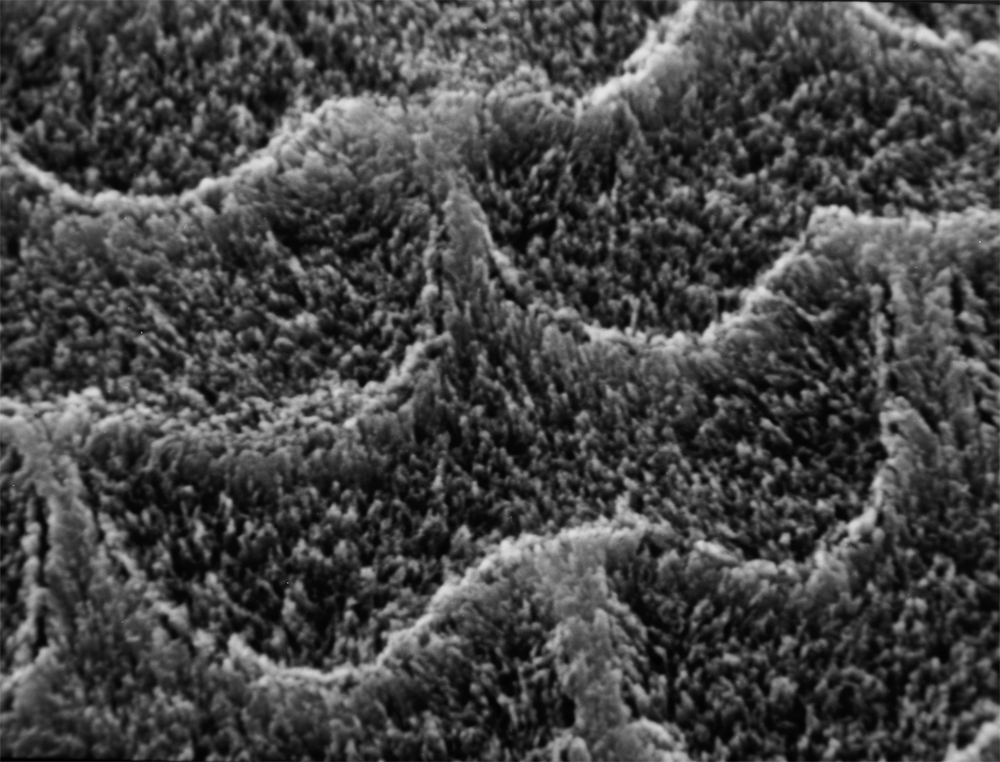 |
| ナウマンゾウのエナメル小柱 |
これとは別に、デスモスティルスという一風変わった動物の歯の研究も与えられました。この動物は北太平洋沿岸のみに生息していた哺乳類の束柱類(目の単位の名称)という仲間の動物で、化石として全身骨格が発掘されていたのですが、生態はもとより、分類上の位置、系統などなどが諸説紛々で、「怪獣」などと新聞紙上で紹介されていたのです。この動物に歯の構造の点から挑戦することになりました。
井尻先生は、ゾウの研究を上記の通り理論的にその特徴を示しましたが、デスモスティルスの研究は「日本的でかつ世界的なテーマである」言われました。日本の独自性が強い動物でかつ世界中の研究者が知っているテーマであるということです(これも私なりに、日本を中心として発見される独特の動物であり、世界中の研究者が注目している、と理解しています)。
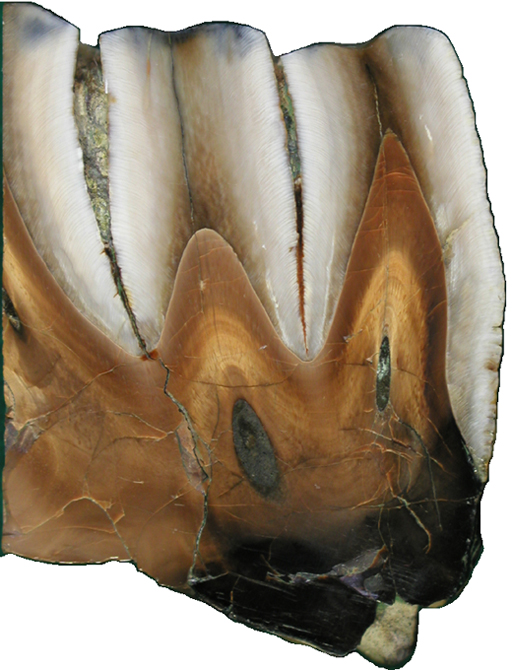 |
 |
| デスモスチィルスの臼歯の断面 白いエナメル質が異常に厚い |
左上デスモスティルスのエナメル質の縦断面、左下が横断面、いずれも光学顕微鏡写真 右上がデスモスティルスのエナメル小柱、右下がデスモスティルスの臼歯 |
以上のテーマはどれも井尻先生のされてきた仕事の一部を引き継いだものです(井尻先生の研究範囲はひろく、文学や哲学、自然科学でも地質学を中心に解剖学や歯学、その発生学、そして実験発生学にまで及んでいます。その歯の一部を引き継いだというわけです)。若気の至りで、これまでの到達点などは度外視して猪突猛進で仕事を始めました。今になって振り返ると冷や汗が出ます。
(反省 残念なことは、井尻先生の歯の発生の移植実験を最後まで引き継げなかったことです。しかし、歯の細胞の近代的な手法を使ってあるていど井尻先生の再生実験を立証できたかな?という感想をもっています)
以上のように三足の草鞋を履いて研究を進めました。周りの優秀な先輩友人ならつゆ知らず、鈍才の私にとっては「メクラ蛇に怖じず」で無我夢中で進めたのです。
そして、長鼻類やデスモスチィルスの歯を解析するためには、ほかの動物の歯を知らなければなりません。そのため歯の構造に関しては約200種弱の観察を行いました。これ以外に歯の発生や分化を検討するために結局、原始的と言われる哺乳類を、さらにさかのぼり、爬虫類や両生類、魚類、果ては火屋(ホヤ)までの歯や石灰化構造に関する形成機構を調べました。その成果が「エナメル質比較組織ノート」と「歯の形態形成原論」です。これで不足した内容を「歯の形態形成原論補遺」としてまとめつつあります。
自分の研究の結果と自己評価
研究の評価は、最初に書いたように、私は自分の研究は基本的に自分で評価するようにしています。むろんできるだけ客観的に評価するのですが、独断てきになりがちです。それを避けるために、自分の三次元的位置をつねに心がけ、他からの評価に耳を傾けるようにしています。
三次元的位置とはよく大学院生や学生に言う言葉なのですが、社会や学会での自分の位置のことです。
例を挙げれば、かつてパスツールが社会的に評価されなかったのですが、100年後の評価はすごいものです。つまり当時の人々にはそれぞれの立ち位置からの評価は下したのでしょうが、普遍的価値についてはだれも気がつかなかったのです。評価とはそのときの社会、あるいは時代の制約があるのです。
ですから、井尻さんの教えのように自分で自分の研究はどのような立ち位置にあるのか考えておかねばならないのです。
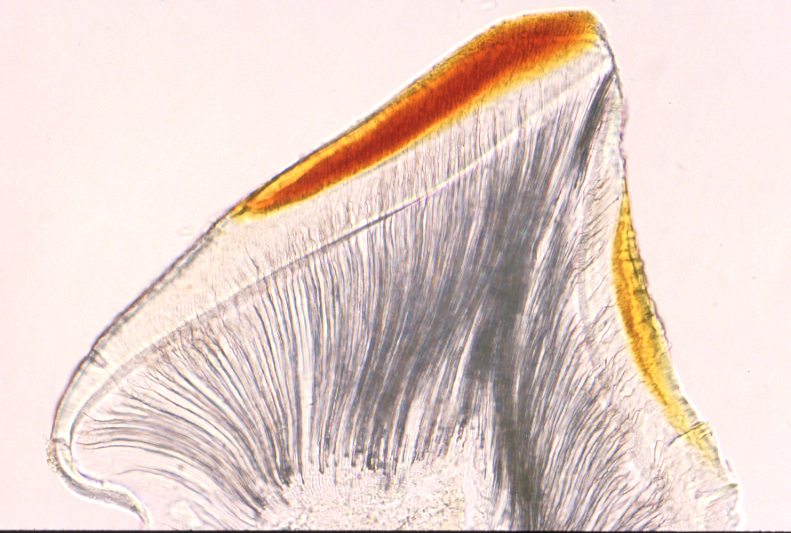
5魚の歯に関して
何といっても最初に手を付けたのが魚(イスズミつまりゴクラクメジナ 図)です。そのわたくしなりの成果は次の通りです。
1 歯堤、歯の原基である歯胚を分化する口腔上皮の間葉への陥入、歯の発生様式は、魚ではものすごく多様であることがわかりました。それゆえに多くの魚類の歯の研究者が新しい歯の形成様式を見つけたという報告があるのです。(最初は、新しい発見が報告されるごとに驚いてばかりいましたが、魚の多様さを理解すると次は何かな?と思えるようになりました。)
(哺乳類では歯堤は歯種ごとにまとまり、(いわゆる永久歯を作る)代生歯堤は必ず舌側(口の中側)へ陥入します。非常に多くの歯のある、しかし歯が交換しない一生歯性のイルカなどもどうもこの様式は同じです。つまり魚の歯の形成様式の一つが哺乳類に反映したものだ、ともいえるのです)。
このような地味な仕事は非常に大切で、多種多様な歯堤陥入の様々な様式が明らかになるのを期待しています。
2 この多様性は、もともと歯は「体の上皮から分化するもの」だからという一般的な回答に、わたくしなりの結論を付け加えています。つまり「上皮の大原則」(上皮の突出に対する対象としての貫入、その様式などだけでなく、上皮の集団化や分泌などの様式)が哺乳類の歯の形成の基本となっている、という考えです(これはあとで触れますが、歯種や歯の交換、歯の構造などの様式の様々な基本原理となっています)。
3 魚類の歯の一番表面はエナメロイドとかエナメル質といわれ、哺乳類とおなじようにハイドロキシアパタイト(水酸化リン酸カルシウム)の大きい結晶でできているのですが、歯の表面としての構造は単純で、膠原繊維などが入っていた痕跡があります。これは完全な上皮性つまり外胚葉性の細胞のみから形成された構造ではなく、かなりの程度中胚葉の関与によって形成されることを示しています。ただし、結晶が大きくなるのは上皮が深く関与することによるもの(私の結晶形成の結果からは、有機基質が上皮によって分解吸収され結晶形成の空間が広くなるため)です。(これに対し両生類、爬虫類、哺乳類のエナメル質はほとんど上皮性の細胞によって形成されます。)
4 よって、しばしば問題となる名称、エナメロイド(enameloidエナメル質様の構造という意味)がよいか、エナメル質が良いかについては、エナメロイドあるいは中胚葉性のエナメル質という妥協案を持っています。結晶がエナメル質のように大きく成長し、硬度もエナメル質とほぼ同じようになるのは、上皮性のエナメル器の組織の働きによる有機基質の脱却(分解し吸収する機能)によることが明らかになっています。
5 象牙質の構造は多様です。骨のような構造の骨様象牙質(osteodentine)、血管が入る脈管象牙質(vasodentine)、石のように構造が見えない岩様象牙質(peterodentine)等など、ほとんど骨と同じような多様性を持っています。それゆえ、これまでの私の研究から、象牙質を「エナメル質ないしエナメロイドと直接接して形成される石灰化構造」と定義できます。換言すれば、上皮に接し直下に形成されるのが象牙質なのです。
(この意味で上皮から離れて形成される石灰化物は、歯の構造に含めるのは妥当ではない、と私は考えています。つまり骨の類なのです。)
魚の歯の研究から「象牙質の定義」をはじめてなしえたことには、ほんのちょっと自負があります。
6 よって象牙質は、基本的に骨と同じ間葉性の構造ですが、上皮からの影響を受けて作られた構造と理解して間違いないでしょう。それゆえに両生類などの象牙質表面、エナメル質との境界には石灰化の高い一層があるのです。このような境界領域の構造的変異は、明確に区分できるものではなく、今後もおおく報告される、というのが私の見方です。
7 魚類では象牙質と顎骨の境界もまた多種多様です。骨に類似した石灰化組織、歯足骨によるばあいもあるし、繊維の場合もあり、また歯足骨と繊維の共存の場合もあります。この構造は明らかに骨なのですが、歯根の周りというより歯根と顎骨の橋渡しのような構造です。これもまた多種多様な骨の構造をしています。また繊維も膠原繊維のみだけではなく弾性繊維などの多様な構造があります。
ただし、爬虫類や哺乳類のセメント質は歯根の象牙質周囲に、言い換えれば象牙質に依存して形成されるのに対し、歯足骨は歯と顎骨の間に独立して形成されます。
次に犬の歯のエナメル質について仕事をしました。このころはエナメル質形成とエナメル質の構造の研究が世界中で盛り上がり、各種のシンポジウムが沢山開かれていました。日本の先輩研究者も前後に落ちず研究の中心的役割をしていました。「Tooth enamel」 「Dental morphology」「Bio-Mineral hard tissue」とうとうの沢山の会議が毎年のように開かれていました。
先輩方が積極的に海外でも発表する姿に非常に刺激を受けました。むろん日本でもひらかれ世界中の研究者が集って、発表し議論しあいました。そのなかにおずおずと参加し、使い慣れない英語で発表し、国内外の先輩諸氏の配慮でどこへいっても議論できるように成長したな~、否、育てていただいた、と感謝しています。
当時、我々若手を可愛がってくださったのは、大学を超えた、世界の恩師の方々であったことも、忘れられません。おもえばグローバルな友情の世界でした。議論も鋭い指摘とともに、温かい指導を含むものでした。相手の弱点を指摘するのを専らとするいまの学会とはちょっとちがう雰囲気でしたね。
さて、イヌの研究では、イヌの歯胚が必要なため、たくさんの仔犬を使わせてもらいました。当時はまだ野良犬がいたためその処理されようとする仔犬を保健所から分けていただいたものです。
当時から今も、ほとんどの研究報告が実験動物として普及していたラットあるいはマウスを使ったものですが、イヌやネコ、ブタ、子牛、サルなどの研究も多少ありました。この少数派は、細胞のレベルではすべて同じだろうというという世界的(今もですが)な考えにたいするアンチテーゼだったのです。そのなかで、どうも種類によっておなじエナメル質を作る細胞(エナメル芽細胞)もちがう様相が浮かび上がってきました。それはよく考えれば当然のことで、エナメル質の硬さも構造も動物それぞれが違うのですから、当然細胞の様相も異なるのです。
この研究の反省点は次の通りです。
1 犬の象牙質表面には膠原繊維の配列がわれわれ人間と違う層がある。これはだんだん動物の種類によってさまざまあることが分かってきました。
2 エナメル質は人間よりもはっきりしたシュレーゲルの条紋という構造を作るが、これに対応するエナメル芽細胞の集団がはっきりと区別できるのです。ここから、エナメル芽細胞の集団と運動の様式の違い、シュレーゲルの条紋の違いが予見されました。その後「エナメル芽細胞の集団と運動」(Ameloblast Grouping and Dancing)として提唱したものです。さらに生物の階層性の関係、体制の原則(体の基本原理)についての理解の端緒でもあり、進化への見方の基礎ができました。
3 この「エナメル芽細胞の集団と運動」(Ameloblast Grouping and Dancing)を象牙質へ応用したのが、いわゆる象牙の縞であり、象牙質のシュレーゲルの条紋であり、かつこの構造はすべての動物の象牙質にあることが予測され、実際ヒトの歯の象牙質でもあることが分かりました。(藤田常太郎の「歯の組織学)ではヒトの象牙質のシュレーゲル条紋を象牙細管の曲走と記載されていますが、シュレーゲルの意図するところは象牙細管の集団の曲走による構造のことでした)。むろんセメント質も形態は異なるのですが、同様の構造があります。
そしていまでは、体のすべての組織、器官に、それぞれ別の名前なのですが、これがあることが分かっています。
4 電子顕微鏡では主にエナメル芽細胞の形態変化、エナメル質の石灰化を観察したのですが、細胞の微細構造の分化のおさらいのようなものでした。しかし、すべてのエナメル芽細胞は模式図のものは抽象化されたもので、すべて異なる形態をすることが分かりました。
ゾウ、そしてデスモスティルスルス
ゾウの歯の研究は医科歯科大学では本来の業務以外の時間、たとえば夜中とかにおこないましたが、埼玉医科大学へうつってから大っぴらに進めることができました。ただし、埼玉医科大学には走査型顕微鏡がないので、師匠の平光先生の援助で日本医科大学で使わせていただくことになりました。
ゾウの系統にそってエナメル質の構造を観察する過程で、対比的にデスモスチルスの歯も観察、それでは十分な比較ができないので、ほかの動物(主に哺乳類)の観察を進めました。エナメル質比較組織ノートの基礎が得られたのはこの時代の成果です。これは今もつづけています。
前にも書きましたが、動物の歯を集めるのに大変苦労しましたが、師匠の井尻先生から譲り受けた標本を中心に多くの方に依頼して少しずつ集めては観察するということが続きました。
1 ゾウのエナメル小柱は、ゾウの先祖といわれるパレオマストドンやメリテリウム(現在は側枝となっている)ではヒトと同じような弧門形(アーチ形)だが、ゴムホテリウムのあたりから銀杏の葉のような形になり、エレファスでは普遍的になること。この形は幅が弧門形よりちょっと大きく、エナメル質の深層(象牙質よりで、組織分化の初期)から生じて徐々に広まること、同時にシュレーゲル条紋は不規則になることが判明しました。
ついでながら、これとともにエナメル質の表面が吸収を受けその表面に歯冠セメント質が覆うようになるのです。エナメル質の厚さもゴンフォテリウムでは8㎜くらいあったのが、エレファスでは2-4㎜ほどになります。同時に咬頭の数が増え、咬頭が横に列を作るというゾウ特有の典型的な形態の歯(臼歯)となります。(但し、エナメル質の厚さを測る基準はありません、そして部位によって厚さが変わるため、ここでいう厚さというのは、全体からの印象のようなものです。決していい加減ではないのですが、比較する基準がないということです。)
これでエナメル小柱とエナメル質、そして歯の形(おもに歯冠)との関係が分かってきました。そしてこの関係は、哺乳類各種ではそれぞれ別の特徴をしめすことも分かったのです。
ちなみに、ゾウのエナメル小柱が銀杏の葉の形であることを報告したのはわたくしが最初ではありません。すでに1924年にJ.H.Mummeryの本にアジアゾウのエナメル小柱は先が凹んだ形をしていると記載されています。これを“maiden hair fern(乙女の髪の毛シダ)”つまり髪が端で膨らんでいる形、直訳としてはシダの葉が並んでいるような形とされています。これではちょっと分かりにくいのでわれわれ日本人によりなじみやすくかつほかの国の人にもよく知られている「銀杏の葉の形」と呼んだわけです。
2 デスモスティルスのエナメル小柱は、小さな楕円形できれいに小柱間エナメル質という壁を挟んで平衡配列をします。一見すると当時盛んに研究されていたラットと似ているのですが、ラットは一本おきに交互に交差して平行になるのです。むしろ、現在のウマに近い形態です。しかしウマなどのエナメル質はせいぜい2㎜の厚さもないのですがデスモスティルスのエナメル質の厚さは8㎜ほどにもなります。このような形態は進化した現在の有蹄類など、植物食に特有なのです。多分きめ細かい構造は強さを増し、植物食を磨り潰す咀嚼に適応している、と想像できます。
わたくしが観察した哺乳類ではこのエナメル質の構造形態と食性の関係は、これ以外の例外はありませんでした。ですから貝を食べるなどの説もありますが、残念ながらいまのところ組織構造と歯の形からみると、常食は植物だということ以外は否定される、という結論になります。
そこで、エナメル質の厚さ、臼歯の形とエナメル質の構造から、デスモスティルスの食性は植物食、それも柔らかい植物であることが言えると考えています。
このように、なにかを証明しようとすると周りの動物との比較が必要なため数多くのエナメル質を観察することになりました。
3 ゾウとデスモスティルスの臼歯の形やエナメル質の構造をみると、一般的に植物を食べる動物は、臼歯形態が複雑になり、エナメル質の構造も規則的なシュレーゲル条と細いエナメル小柱の規則的配列、ということになります。そしてこの細いエナメル小柱をつくるのは細長いエナメル小柱であることも明らかになりました。ちなみに肉食動物のエナメル小柱はアーチ形か円形または六角形の横断形と幅が広いという特徴があります。
これらのことは、植物を食べるときはするつぶすための咀嚼が行われるため、エナメル質を含む歯冠がすり減るのに対する抵抗するため、高い歯冠とエナメル質構造が複雑に規則的になったと理解することができます。(目的論のようですが、そのような変異が適したという意味です)
ただしゾウの系統ではエナメル小柱が組み合わさったシュレーゲル条は不規則な方向へ進化し、デスモスチィルスでは規則的な方向へ進化します。デスモスティルスと同じような変化をするのはウマの系統で、サイなどはまた違った方向へ進化します。
しかし、これは一般的な話で、これ以外にも例外が沢山あること、つまり適応形態は多様でありそれぞれ最適な形となっていることが、比較解剖学的な研究からわかってきました。私はこれを環境にも種の嗜好性(固有性)にも依存ずる安定的、平衡的状態への変化と考え、体制の原則にしています。
*嗜好性という用語については、和気先生から意思性が入るので、細胞などに使うのはどうか、という貴重なご意見をいただきました。細胞や組織、器官では親和性などが使われますがピッタリする用語がないのが現状です。むしろ「好み」と単純に表すのが良いのかもしれません。やさしい誰にでも意味がとれる用語があれば、またそのほかのことでもご意見をいただければ幸甚です。
4 埼玉医科大学時代はわずか2年でしたが、エナメル質の比較解剖の仕事が進み、数十種類の動物の進化の様子が分かりました。
その一つが上記のウマの系統です。ウマの系統は、進化に従ってエナメル小柱が幅5μくらいの弧門形から幅2μくらいの楕円形へ、互い違い配列から平衡並列へ、しかもエナメル小柱の各列の間には小柱間エナメル質という層を挟むのです。シュレーゲル条も規則的に進化します。ほぼデスモスチィルスと同じです。
ただし、象牙質は違います。原始的なヒラコテイリウムやメリキップスでは我々人間と同じ組織だったのですが、メゾヒップスあたりから象牙細管が太くなり、いまのウマではものすごく太くなるのです。この細管の中は二次的に石灰化するので細管の中の象牙質を管周象牙質とよびます。つまりメゾヒップスあたりから管周象牙質が厚く発達するのです。これは歯冠の高さが増す時期と一致しているので象牙質の嵩が増えることへの適応と考えられます。このような進化は長鼻類にも認められるのですが、ウマほど著しいものではありません。まあウマの仲間特有といってよいとおもいます。ここからも適応はそれぞれの動物で特有であることが分かります。
ただ、サイの系統のエナメル質は各種類の特徴がばらばらでまとまりません。シュレーゲル条も非常に特徴的で捉えるのに苦労しました。ふつうは歯冠の横方向に違う模様が配列する、つまり縦に切ると模様が見えてくる、のに、サイの系統では縦の方向に模様、歯冠を横断すると模様がみえる、のです。これは歯の形成方向から考えると非常に難しいのですが、グルーピングの様式が違うことで、理解することができました。
まずもって、種の特異性を認識されたということです。ここから前にふれた種の特異性を実現する嗜好性が体制の原則であること、進化に関与していることを確信しました。
日本大学へ移ってからは、スタッフの充実したこともあり、歯の発生の仕事を広めました。まず人間の歯に類似するといわれたブタ、それに大学院生の研究していたウサギの知見も加えることができました、さらに、イギリスのマンチェスター大学でポッサム(有袋類の袋ネズミともいう)をすべて廃棄するというので譲り受け、原始的特徴を持つという有袋類、さらに食虫類、ワニやイグアナなどの爬虫類、両生類はスタッフが研究していたアフリカツメガエルの研究から勉強し、教室の先輩から火屋を試料としてもらい研究に加えました。また大学院生のアメリカザリガニの研究から、歯とは何かを考えるきっかけをつかみました。
その過程で大学院生や臨床教室からの研究者が臨床的ないろいろの研究をしているので、このような比較解剖に役立てさせてもらいました。たとえば、エナメル質の結晶がどうして大きく成長するのかについて、結晶の初期形成(結晶核形成)と結晶成長について、化石の結晶を観察し、イヌやブタの結晶形成とつなぎ合わせ独自の理論まで高めることができました。これは歯の形態形成原論に書いてありますが、結晶のみ、あるいは細胞の研究では得られなかったものと自負しています。その一方、両方ともに研究する人はすくなく理解されにくいのだろうな、と感じています。
しかし、大学院生をはじめスタッフの方々に感謝しています。歯に関する主なものを挙げると次のようなものです。(歯以外では血管の新生、骨の代替剤、神経の再生など、沢山の仕事を経験させていただきました。)
イヌの研究、ブタの研究、ラット、マウスの研究、ウサギの研究、有袋類のオポッサムの研究、食虫類のスンクスの研究、爬虫類のワニの研究、イグアナの研究、両生類のアフリカツメガエルの研究、脊椎動物の起源と言われるホヤの研究、甲殻類のアメリカザリガニの研究(殻と石灰化の問題)
ヒトの胎児や幼若個体における歯の形態形成の研究
*ヒトの歯の研究にはとくに新潟大学の歯学部解剖に保存されていた標本、鶴見大学の解剖学教室の標本、そして私個人の標本をつかいました。雪がないという新潟市に大雪が降り、何日もμCTを使わせていただいた日々が昨日のようです。本当にこれらの大学の方々にはお世話になり感謝の気持ちでいっぱいです。あとで述べますが、このようなヒトの研究では動物で得られないデータを得ることができ、あとで反省するようにヒトの歯や進化の考えを一層推し進めることができたということを記しておきたいと思います。それはなぜかというと、ヒトの歯のデータは動物に比べて比較にならないほど蓄積されているからです。この蓄積のうえに自分のデータを照らし合わせて進んだという、幸運です。
このうち歯の発生の免疫組織学的ないし細胞学的研究は主にイヌの歯胚を使いました。
この成果は歯の形態形成原論に載せていますが、そこで得た大切な点のみ振り返っておきます。
イヌの歯のエナメル質には非常に明瞭なシュレーゲル条があり、それに伴う運動が免疫組織学によって組織にも細胞にもあること、これを前に既述したように組織細胞のGrouping and Dancing(集団化と動き)としましたが、それには周期性(律動)があることが証明できました。さらに重要なのは、電子顕微鏡像からはトームス突起の中の細胞小器官の変化から推定していたのですが、エナメル芽細胞の分泌周期を明らかにできたことです。
これには驚いたことに、エナメル芽細胞の動きを咬頭全体から眺めると、咬頭頂を取り巻くように、2種類のエナメル芽細胞の集団が渦巻き状に配列しているのです。まるで星雲です。(このようなこともあり、自然と体の細胞の関係についての「体制の原則」に確信を持ったのです。まあ当然といえば当然なのですが・・・)
 |
 |
| エナメルが細胞の群れが咬頭を中心に渦状に配列、 咬頭見た写真 |
NASA公開画像 M51 Hubbleによる星雲 |
以上の集団化は進化や発生過程の分節化と連動し、周期性は多様性がありながらも生物に共通の特徴であることに発展させることができました。だから、これらは体制の原則として一般化したのです。
エナメル芽細胞にも形態的特徴があり、大きく分けると食肉類にみられるような細胞小器官の分散傾向とラットなどにみられる組織的な規則的配置傾向があり、前者の細胞の丈は短く、後者の丈は長い傾向があります。つまりラット型は細く長く細胞小器官は規則的に組織的配列を示し、食肉類型はずんぐりむっくりで、分散的不規則的な細胞小器官の配列をしているのです。
この二つのタイプとエナメル質の組織を関連させて考察すると、ラット型が楕円形のエナメル小柱が小型で平衡配列かそれに近い型で、食肉類型は円形あるいは弧門形のエナメル小柱が斜め配列か不規則に配列しているタイプです。どちらもシュレーゲル条は顕著な場合と目立たない場合があります。
これと歯の形は直接結びつかないのですが、ラット型は有蹄類などの植物食で歯が長く、エナメル質が厚い場合が多く、食肉類型は歯が短く歯根が形成される場合が多いようです。
ただし、エナメル質の厚さの比較は正確にできないのです。しかし、全体的に歯を見ると厚いエナメル質と薄いエナメル質があります。極端に厚い例は、ゾウの先祖やデスモスティルスなどで6-8㎜もあります。薄いのはクジラなどで1㎜もありません。これは繰り返しますが、全体的に大まかにみて、ということです。
ラットと同じ無根歯といわれるウサギの切歯や臼歯では、シュレーゲル条はやや有蹄類型と食肉類型の中間ですが、弧門形から楕円形のエナメル小柱が平行ないし交互配列をします。エナメル芽細胞も食肉類型とラット型の中間です。どうもエナメル質形成の速度がラットなどよりゆっくりのようです。
さてラット型の典型はブタで、楕円形の小型のエナメル小柱が平衡配列します。この様子からエナメル質形成の速度と細胞の形態が連動しているようです。食肉類型はネコやイヌなどで、歯根がしっかりと形成され、歯の形成時期が限られているため、エナメル質形成速度が比較的ゆっくりの印象があります。
しかし、これにもたくさん例外があり、ヒトでは、弧門形のエナメル小柱が斜めに配列し、ゆっくりと形成され、エナメル芽細胞は食肉類型です。インドゾウやアフリカゾウでは、弧門形から銀杏の葉型の比較的大型のエナメル小柱で、食肉類型のエナメル小柱を予想させますが、エナメル質は厚く、形成速度もそれほどゆっくりとは考えられません。そして植物食です。反対に、ウマやウシでは(それほど沢山観察はできませんでしたが)、ラット型のエナメル芽細胞と小型の楕円形のエナメル小柱が平衡並列するというもので、エナメル質形成速度も速いようです。
このようしてみると、食性や歯の形態などでエナメル質を分類するには非常に多様であることが見えてきました。つまりそれぞれの動物なりに適応しているのです。それぞれの動物にとってその適応が妥当ではないだろうという印象をうけるものもあります。
このようなことをもとに導き出したのが、種の特異性は嗜好性にかなり依存している、ということです。多様性は嗜好性に依存している面が多い、というものです。多様性のある適応は、必ずしも最短、最適のものばかりではなく、それぞれの種の嗜好性も加わって選択するのです。これでかなりの生物の現象、そして進化の現象が腑に落ちるようになりました。まさに生物は「蓼食う虫も好き好き」なのです。
以上のような歯の研究の進行とともに、有袋類などに代表されるエナメル細管の起源、歯の形態進化、歯列の形成(歯数、歯種はどのように決まるのか)、顎とは体におけるどのような意味を持つのか、などなどの疑問を研究し、その成果が「歯の形態形成原論」となりました。この本は私の名前で出版しましたが、半分は一緒に研究した方々の成果です。
8 結晶の研究
結晶の研究も私にははなはだ大変だったのですが、よき師よき先輩に囲まれ、いろいろと勉強することができました。
主な成果は次のようなものです
結晶核(結晶の種、最初の結晶の先駆物、などとも表現されている)は、エナメルタンパク質のマイクロスペースで形成される。(マイクロスペース説)
結晶成長は単結晶の大型化だけではなく癒合もある。
象牙質の結晶は平板状であることを直接観察。
エナメル質の着色(ラットの切歯エナメル質の表面、トガリネズミのエナメル質の表層半など)は鉄の沈着であるが、鉄分子が結晶格子不正と結晶の表面であることを立証した。
エナメル質の結晶は六角柱であるが、これを原子間力顕微鏡(AFM)などで観察し、結晶は癒合なのにより必ずしも六角柱状ではないことを明らかにした。
研究の環境
東京医科歯科大学の時代はエナメル質とエナメル質を作るエナメル芽細胞の研究が中心でした、それは解剖学教室では比較解剖学とともにエナメル質形成の研究に重点が置かれていたためです。エナメル質は体の中でも大きく目立つ結晶で、これを細胞がどのように作るかが一つの焦点になっていたのです。とうぜん我々若手もその波に呑まれていました。
しかし、生体鉱物とか生体結晶といいながらも、細胞と結晶の溝は深く、両者を同時に理解するのは困難を極めました。この問題は、いわゆる境界領域であり、学際領域の問題として、いっとき話題になりましたが、この状態は今でもさほど変わっていないようです。
いっぽう私は、解剖学者でありながらエナメル質の結晶を研究し、結晶学に造詣の深い平井先生、元来は形態学でありながら結晶形態を極めた一条先生等にかこまれ、さらに留学先は鉱物学出身でありながらエナメル質の結晶とその形成を研究しているHöling先生であり、結晶学の碩学に囲まれるという幸運にもめぐまれた。このような境界にあって、ほかの人達よりほんのわずかに両領域を同時にふかく理解することができた、と感じています。その結果が上記のものです。
いずれも世界でほぼ初めてのものなのですでが、物理学や化学の領域の成果を根拠にして間違いはないだろう、と自負しています。ただし、理解してくれる人は非常に少ないので、やはり学際領域なのです。
ここで「ほぼ」というのは、マイクロスペース説は、私がエナメル質形成の超微細構造の観察から見出したものですが、象牙質では生化学的な研究からすでに仮説(ナノスペース説)がたてられていたのです。
余談ですが、この研究者に私の成果の是非を確認した折、海外の雑誌に投稿するのはよした方がいい、査読中に盗まれた、と助言をいただいた。げに、さもしい世の中なのだ、と感じました。
象牙質の六角形の平板状結晶は、透過型電子顕微鏡による結晶の断面の観察と電子線回折などで予想されていたものでしたが、結晶の癒合による成長と共に、全体の姿がスッキリ観察できたのはまったくの幸運で、海の底にあった象牙の有機質のみがきれいに洗い流されて結晶だけが残った結果です。
それは結晶だけではなく、なんと長年夢見てきた象牙質の景観も提供してくれたのです。次の写真を味わってください。なんという自然の力でしょう。
 |
 |
| 象牙質の結晶が六角の平板状であることがわかる | 中央の象牙細管の中の様子、結晶の配列から膠原線維の配列までわかる |
それまでは、試料として使う象牙質は膠原繊維と結晶が密な構造を作っているもので、コラゲナーゼという膠原繊維を溶かす薬剤で処理する方法をいろいろ試したのですが、純粋に結晶だけにするのは困難だったのです。
結晶の研究過程を振り返ると・・・
エナメル質、エナメル質形成の研究をおもに光学および透過型と捜査型電子顕微鏡でおこなったため、試料は薄切して染色し、その断面を観察するという方法によりました。薄切とはいうがこれは試料を壊すことだし、染色によって(酸を使うため)当然アパタイトの結晶は溶失してしまうのです。そのため結晶の薄切面をじかに電子顕微鏡で観察する方法がとりました。
当時はガラスナイフによる切片作成が主体で、ようやく高価な、それも超高価なダイアモンドナイフが出始めたころです。ダイヤモンドナイフを科学研究費で買えたのは1980年代になってからです。エナメル質は固く(モースの硬度計で6~7)、ダイヤモンドナイフを、唯一回の薄切操作で破壊したことが何度もあります。
特に化石のエナメル質は硬く、化石のデスモスティルスのエナメル質の透過型電子顕微鏡で観察した論文が出たとき、何十本もこの超高価なダイヤモンドナイフを使った、と記載されていたのには驚きました。まさにアメリカと日本の科学経済状況の違いを目の当たりにして、ため息と共にいつかこれを成し遂げてやろうという気持ちを持ちました。
ただ師匠の一条先生のみは、一本のダイヤモンドを駆逐してこの超薄切片に挑んでいました。そして、世界で初めてエナメル質の結晶の原子と原子配列を観察した、まさに鬼神の技なのです。
私の感覚では、人間のエナメル質がもっとも硬く、ついで化石動物、ラットやトガリネズミは柔らかいエナメル質という印象でした。ちなみに当時公表されていたほとんどの研究はラットのエナメル質です。そんなこんなで、エナメル質の薄片(電子顕微鏡で観察するため40-70㎚の厚さの薄切片をつくる)が切れたのはドイツに留学した時でした。
銀色の薄切片がスパッと眼下で切れたとき、なんと嬉しかったことか・・・。それは、電子顕微鏡用の切片を作り始めてから十数年も経ていたのです。とにかく諦めずに続けることの大切さを学んだのです。
ちなみに、原子配列を観察するためにはできるだけ薄くする、切片の厚さは溶液の上に浮かんだ切片の反射光の色(波長)から判断する、できるだけ薄い銀色(40nmくらい)のような状態まで薄くするのです。帰国して師匠の一条先生を訪ねると、相変わらずエナメル質の超薄切片を作成しており、「小澤君、切片が見えるか?」と質問された。薄切機(ウルトラミクロトーム)をのぞいても見えないのです、あれ?「君、水の上のごみを見なさい。動くだろう、それで切片が切れているのを判断するのだ」と諭された。確かにナイフの動きに沿ってゴミが動いている。げに、超人的技術なのです。
一条先生の思い出にはこんなものもありました、学会で「どうすればそのような切片が切れるのか?」との質問をうけ、「厚くもなく、薄くもなく、早くもなく、遅くもないように切るのです」と答えていました。まさにその通りであり、これはエナメル質を切ることができる、それもヒトなどの硬いエナメル質を着ることができる人、のみにしかわからないかもしれない。
さて私の超薄切片をつくることができた原因をいろいろしらべ、ドイツの寒冷な、そして乾燥環境での薄切がよかったという結論に達したのです。さっそく帰ってから薄切室に乾燥設備を設置した。ダイヤモンドナイフを買うことも、この設備も、研究費もなければできない相談で、科研費をはじめとする研究費をいただいたことに感謝してもしきれない思いです。研究は一人ではないということを実感しました。
このころは鉄がエナメル質を構成する結晶のどこに入るのか、入る機構はどのようなものか、という興味が中心で、それゆえにドイツのHöhling教授(ミュンスター大学医学物理学研究所)のもとに留学したのです。教室の総力を挙げた仕事になり、レーザー微少質量分析器LAMMA(当時富士フィルムにしかなく、ドイツで使わせてもらいました、ちなみにエナメル質に応用するのは我々が世界初でした)、エックス線回折、示差熱分析、赤外線吸収、電子線回折、電子顕微鏡による原子配列の観察、とうとうを駆逐した研究でした。結果は、結晶格子不正の位置、結晶の周囲などに鉄分子があることが分かりました。エナメル質形成過程で鉄分子が沢山あるためこのような結果になる、のです。
結晶格子の研究は、結晶格子不正がトガリネズミで沢山観察されるところから始まってこのような結果になったのです。
ちなみに、鉄を含むエナメル質と含まないエナメル質の境は破損することが多く(下図)、
 |
 |
 |
| トガリネズミの歯の断面 象牙細管、エナメル細管という管がはしる 上の方の茶色の領域が鉄を含むエナメル質 |
エナメル質の徴薄切片 鉄を含む上と含まない下の間が壊れる |
エナメル質を酸でほんのちょっと溶かすと、鉄を含む右の領域は残り、酸を含まない左の領域は溶ける |
エナメル質に鉄がはいると、硬いうえに、粘り強くなる、しかも酸に対して抵抗性がある、ということが薄切の過程からなどの印象です。ラットなどが木を齧るときエナメル質が破壊されない効果があるようです。
つぎに化石の結晶の原子配列の観察に移ったのですが、これも化石は何千万年も経たものもあり切片を切るのに困難を極めました。その結果がデスモスチィルスやマストドンなどの結晶の格子構造である。ここではこれらの化石の結晶格子が形態的には化石化による影響をあまり受けていないことが判明しました。そこから、上記の象牙の結晶の観察が二次的な化石化の影響をそれほど受けていないであろうと、考えています。
つぎに化石の結晶の原子配列の観察に移ったのですが、これも化石は何千万年も経たものもあり切片を切るのに困難を極めました。その結果がデスモスチィルスやマストドンなどの結晶の格子構造です。ここではこれらの化石の結晶格子が形態的には化石化による影響をあまり受けていないことが判明しました。そこから、上記の象牙質の結晶の観察が二次的な化石化の影響をそれほど受けていないであろうと、考えています。
ハイドロキシアパタイト 六角柱の断面探し
ヒトのエナメル質を構成する結晶、即ちハイドロキシアパタイトの結晶は自然に出来る結晶が六角柱であること、ラットの未完成のエナメル質、つまり有機基質がすごく多い時期の結晶を、超音波処理などを施して有機基質を取り除いた結晶が六角柱の針のような形をしているため、などから六角柱であることが推定されました。このことから、六角の断面から原子配列を観察するのが一番よいということで、エナメル質の薄切切片上から六角の結晶探しが始まったのです。ラットなどでは、比較的容易に六角の結晶断面を観察できるのですが、ヒトではこれが極めて難しいのです。切片を舐め尽くすように隅から隅まで、電子顕微鏡で観察しても六角の断面は容易に出てきません。なぜか?しかし、六角の断面でなくとも結晶格子は観察できるのです。
結晶形成の過程から考えて、大小様々の六角の結晶が癒合しているのではないか、と推定しました。つまり、結晶形成過程の動物による違いは、有機基質の脱却様式の違いがあり、この違いは結晶の場であるマイクロスペースの違いとなり、よってヒトでは大小様々な結晶が癒合する、と考えられるのです。ラットなどでは、エナメル質形成の速度が速く結晶形成の場が比較的早く大きくなるのではないかと推定されるのです。
つまり、ヒトのエナメル質に限らず動物のハイドロキシアパタイトは有機基質がベースとなって形成される、よって自然界のハイドロキシアパタイトより不完全な結晶だろうと、予測し、期待していました。そこで、我々の扱っている結晶は自然界の結晶とどのような違い、弱点?がああるのか、鉱物学の権威で電子顕微鏡による結晶格子を観察している先輩(京都大学理学部地層鉱物学の富田さん)に、おそるおそるヒトなどのエナメル質の結晶格子の電子顕微鏡写真を見ていただきました。
結果は、予想に反して「よい結晶だね~」とのこと。我々にとっては期待外れ、ガックリの一言でした。要するに結晶自体には動物も自然界の結晶と差はなく、結晶形成過程からくる結晶の形の違いや鉄などの沈着による違いがあるのみなのです。
まあ、当然と言えば当然のことなのです。
ここから動物間の結晶の違いが推定されました。たとえば、ゾウの牙などでは、ラットと同じように長細い六角形の断面が観察されます。
こうして透過型電子顕微鏡による結晶格子の観察は一応終了しました。
結晶観察の反省
これはすべての観察に通じることなのですが、このような結晶の電子顕微鏡などによる観察は、観察できるようにする準備過程で試料に様々な処置を施します。よって見えるようにする過程での試料の変化も考慮に入れなければ何を見ているかわからないことになります。また電子顕微鏡では試料にじかに電子線を充てるためにその影響もあります。なによりも薄切による二次元的な観察が主体なので、それを三次元に組み立てなければなりません。そのうえ、化石の試料では化石化の過程での変化も考慮に入れなければならないのです。よってなにを観察しているのか、できたのか、十分な反省が必要になってくるわけです。これは我々形態学を専攻する者にとって逃れられない宿命です。
(私は、体の解剖学的分析から、ここで最も大切な「感性」はおもに幼少期に涵養されること、それには野外での遊びが必要であることを導き出しました。ちなみに、親が忙しく、子供をふりまわしたり、ジャレたりすることが少なくなった今、特に都市部では終日の公園等を使ったあそびが独創性につながるのです。)
おおよその経過
師匠の井尻さんから「仕事を纏めるよう」再三言われていましたが、定年まで10年を切る頃からやっと本にまとめ始めました。反省をこめて振り返ると、仕事のまとめを進めることで研究者になる最初の目的に立ち戻った、と感じています。当初、井尻さんに入門したとき「進化の研究をしたい」と青臭い希望を述べたのですが、井尻さんから「進化を研究するために、まず解剖をまなぶこと」と命じられ、解剖学や発生学、歯学を学びつつ、与えられた研究テーマを皮切りに仕事をすすめ、これを纏める段階で、進化の問題に立ち戻ったのです。その経緯は次の通りです。
まとめは大きく分けて3期になります。まず、生のデータをまとめる、ついで、そのデータから法則を抽出する、最後に、進化の問題を振り返る、というものでした。進化の問題は今進行中です。
このまとめと並行して、現在、歯学部や古生物学等々で広く使われている歯の法則の原典を翻訳しています。
今使われている歯の法則は「三結節説」と呼ばれ、アメリカの古生物学者のCope-Osborn-Gregoryという師弟が提唱した考えです。全世界に広く普及しています。しかし、これには、今の研究者は(少なくとも日本では)原典に触れていない、そして少なからずこの学説には瑕疵があるのです。それは、どれほど膨大な標本を観察して抽出した法則とは言え、百年以上前に書かれたものですから当然のことです。その問題の一つは、臼歯の歯冠のみの理論であること、例外が少なからずあることなどです。
このような理由以外に、解剖の先輩が海外の優れた解剖学書を翻訳してホームページへ載せているのを知ったことや、かのW.VOGTの「両生類胚における造形運動と器官形成」を波磨忠雄氏などが仲間と十年以上をかけて翻訳したことが刺激となりました。この本は絶版になっているのですが、発生学の原点とも言えるものなのです。翻訳に関すれば、私の師匠の一人、萬年甫先生が、かのラモニ・カハールに関する「神経学の源流、ラモニ・カハール」を訳されたことも大きな刺激になりました。カハール染色は神経を染め出すには避けて通れないものだからです。
私の領域は歯という狭いものですが、秀才の脇田さん(北海道大教授名誉教授)が歯列の形成機序の著作、私がCope-Osborn-Gregoryと、分担して進めています。是非、今最先端の分子生物学や遺伝学で歯を紐解いている若い研究者に目と通してほしいものです。
さて話を私の研究のまとめに話を戻すと、仕事の第一弾が「エナメル質比較組織ノート」です。第二弾がデータと法則の抽出をおこなった「歯の形態形成原論」です。仕事の内容から「原論」という題は恥ずかしかったのですが、歯の研究の世界を見わたすととこう言ってもよいかなという感じを持ったのです。日本はもとより世界でも歯の形態全体を論ずることが少なくなっていたからです。
第三弾は、いま書いている「歯の形態形成原論補遺」です。歯の形の基本的な問題、進化の問題まで踏み込むことになります。前の2冊は、出版社と印刷所、教室の方がた、そして出版助成金を出してくださった大学、に大変な援助を受け、心から感謝しています。
ただし、補遺の仕事は、本当に大変な仕事になりました。これまでにほとんど手をつけられていない歯の問題、そこから派生する様々な問題が次から次へと現れ、当初2−3年と予定していたものが、十年を過ぎた今も終わりがぼんやりとしか見えないのです。
これ以外に「私の解剖学原論」を書き始めています。
解剖学の専門書は、すぐれたものが多いのですが、どうもその心、つまり哲学的な部分が不足しているように思えるのです。例えば、人体の階層性として、体ー器官系ー器官ー組織ー細胞をあげる教科書が多いのですが、この意味は書かれていません。意味は殆ど自分で考えろ、というのでしょうか。
その結果、階層性は体の構造を整理したもの、と考えているかたが殆どだと思います。そうなのですが、階層性は独自の生きる法則を持つこと、各階層間は連絡調整があることなどが、やはり書かれるべきだと感じています。
この一連のことから感じることを一言、言わせていただければ、昨今は(かつて自分もそうだったのですが)教科書を書くことで、自分の研究のまとめをしない傾向があります。これは残念なことで、定年が見えてきたら仕事のまとめをする風習が日本に定着することを願っています。なにより知識の財産を残すこと、自分の仕事から抽出する法則を後輩に伝えること、が大切だと考えるからです。
出版
出版の問題ですが、民間の出版社では「売れる」ことがないと困難なので、大学ごとに出版局をつくるなどの努力があればよいなと感じています。簡便な方策としては、出版補助金をだすのもよいでしょう。いろいろな助成金があるにはあるのですが、まだまだ十分でなく、知識が埋もれてしまうのは大きな損失です。私もその例に漏れず、色々方策を練っているところですが、一つには自分のホームページを作ってここに発表するようにしています、次に、今流行りのネット出版も良いかな、と考えています。
さて、仕事をまとめて感じることは、まだまだ自分の結果には穴があることが分かり、補遺に挑戦する気持ちを改めて固めています、しかし、最初からの問題意識は同じで、ここから進化の問題まで踏み込んでいます。かつて大学を出て、井尻さんに「進化」の仕事をしたい、という原点にもどったことを、幸運だなと感じています。
生涯の疑問
学生時代の最初から抱いてきた疑問を解説しておきます。一言で言えば階層性の関係の問題なのです。
これまでそして今も、それぞれの階層ごとに詳細な研究がなされ成書に書かれていますが、相互の関連性についてはほとんど記述されていないのです。一例をあげれば、歯のエナメル小柱とシュレーゲル条の関係、シュレーゲル条と歯冠の形態の関係など、です。これらは有る特定の動物では、関連性があるように書かれていますが、他の動物ではまた違った関連性を持つために、相互に関連することは確かなようですが、どのような関係にあるのか、普遍的な関連性があるのかないのか・・・、という疑問です。
ちょっと詳しくのべれば、エナメル小柱が小型化し規則的な配列をするのは有蹄類など植物食特有のもののようですが、植物食でありながらエナメル小柱が大型となる長鼻類がいるのです。このような疑問は研究が進むにつれて大きく膨らみました。このような例外をどのように説明すればよいか、いつも胸に抱いていました。
そして、この疑問は仕事を纏めることによって一定の回答が得られるとともに、生物(生命)の進化の問題がくっきりと浮かび上がり、生物(生命)の進化の理解がすすみました。これが、ホームページの「進化」の「進化理論と進化の定義」に要約してあります。
最後に一つだけ問題点を挙げておきます。
古生物学関係の方が理解しにくいのは、個体発生と系統発生の関係のようです。ぜひ理解してもらえるととてもありがたい、と考えております。もう一つ苦手なのは、突然変異の捉え方です。生物の代謝や遺伝は無数の変異があり、目立つ突然変異が進化に結びつくには、どのような過程を考えなければならないか、ということです。
一方、現生の生物を扱っていると、地質学的年代の捉え方がどうも苦手なようです。古生物学で突然、といえば明日の朝にも・・・という時間の感覚になりますが、万年単位の突然を捉える努力が必要です。
つまり、時間の捉え方が両者では全く違い、ひいては学説を理解できないことにつながっているようです。
もっとも残念なことは、ここまでたどり着いた時にすでに師匠の井尻さんは幽界に旅立たれた後であったことです。歯の形態形成原論の完成前には、師匠の一人大森昌衛先生に原稿を見ていただき、前書きをいただきました。また、医科歯科大学時代の萬年甫先生も同様です。これには、一条先生と同期の本松清行先生に大変お世話になりました。なんとか間に合ったかなという思いです。しかしその後相次いで幽界に旅立たれ、さみしい限りです。
まえにも書いたのですが、とくに井尻先生とは、歯の形態形成の原理、個体発生と系統発生、これを結ぶ体制の原則、そして自然弁証法について一番議論を交わしたかったのです。なにはともあれ、古生物学も現生の生物学も、哲学も、同時に理解する先生は他に見当たりません。私の師匠であり、比較解剖学の泰斗のあの三木成夫先生にしても、古生物学の重要性は十分認識していたものの、そこまで手が届いていなかったようです。
そして、今の大学(世界の状況も同じですが)ですが、論文数で競う成果主義あるいは論文の掲載雑誌による評価主義(インパクトファクター)に対して、自分が十分に抵抗し得なかったかな、と反省しています。やはり、研究は上に書いたように、基準あるいは視点をしっかりともって、基本的には自分で評価するしかないのです。評価は人によって、時代によって様々に変わります。まず絶対といえる客観的な評価はあり得ないのです。
この点で、自分の関係した大学院生には(あるいは論文博士の方にも)、とくに主論文は、自分の言いたいことを自分のデータをもとに日本語でもいいから全て記述するようにと、最後の一人まで指導できたのは、よかったかな、と感じています。