
爬虫類の歯
爬虫類は中生代に地球上に最も繁栄したとされる動物です。水棲と陸棲、水棲は海棲と淡水棲がいます。そのご大形の爬虫類を中心に消滅しますが、今なお哺乳類よりも種数は上まわっており、鳥類と爬虫類を合算すると哺乳類の3倍くらいの種数となります。ですから歯と言っても多種多様ですが、爬虫類の歯の特徴は、一口で言えば、「哺乳類の歯の多様な発達の準備段階」と言えるようです。その主な特徴は次の通りです。
1 歯は顎の骨、上顎は前顎骨(切歯骨)と上顎骨、下顎は歯骨(下顎骨)に植立する。ただし上顎も下顎も複数の骨からできています。(哺乳類では歯のある骨、つまり上顎は上顎骨と切歯骨、下顎は下顎骨からできています。)
この顎の分節に相当する一群の歯をJW Osborn(1984)は「Clade(歯族)」と呼んでいます。顎の分節の歯とは明言していないのですが、意味はほぼ同様です。しかしOsbornは分節が哺乳類に向かって増えると考えました。((哺乳類は切歯、犬歯、小臼歯、大臼歯、あるいは乳切歯、乳犬歯、乳臼歯という歯種があります。注;歯種というのは一般的な用語でこれ以外の区分にも使われます。))
つまり爬虫類の特徴は顎の分節がハッキリするという特徴があるのです。
| |
クチヒロカイマンの歯列 歯の大きさが変化し何回か繰り返している。 |
4 歯槽に生えている歯は、その下、舌側がわから次世代の歯が生え、歯髄のなかにはいってきてしまいます。(哺乳類の乳歯と永久歯の交換様式とおなじですが、次世代の歯が歯髄のなかに入ることはありません。)
 モササウルスの歯の交換 機能歯の下に次の世代の歯がある
モササウルスの歯の交換 機能歯の下に次の世代の歯がある
 |
 |
左:ワニの歯の交換 舌側からのみたところ、歯の中央に縦に浅い溝が走り、舌側の根尖が開いている。そこから次世代の歯が覗蹴る。 右:下顎断面:機能している植立歯は歯根膜で顎骨と結合し、左下に次の世代の歯胚がある。丸いのはメッケル軟骨と神経線維側 |
5 歯の交換数は多生歯性です。(残念ながら、哺乳類の二生歯性と爬虫類の多生歯性の中間型はみとめられていない、というのが現状です。)
爬虫類は体が大きく成長するにつれて顎も歯も大型となりかつ歯の数が顎の奥に向かって(遠心側へ)増えます。顎は奥の方に成長するので、歯も奥の方へ奥の方へと増してゆくと言うわけです。歯の交換する回数はどのくらいかと言う点はあまり分かっていませんが、終生生え代わるようです。ワニの歯の交換数を数えた方が約60回だったと報告しています。
しかし、歯が交換するまでの期間(歯の形成から脱落するまでの期間)は、当然ながら歯が小さいほど早く、成長して体と歯が大きくなるにつれて長くなるようです。ワニの歯の例をとっても、最初の歯は一ミリも満たない大きさですが、成長すると数センチの長さになります。これを裏返せば周期性は年齢とともに変わることがわかります。
さて萌出し機能した歯は一定の時期が来ると抜け落ちます。これは後から生える歯(後継歯)によって押し出されるのではなく、ある交換の時期が来ると歯髄や歯の周囲に破骨細胞が現れて歯を溶かしてしまい、残った歯が根本から折れて脱けおちます(隣にある歯の吸収はしないので歯の萌出の周期性といえます)。
抜け落ちるとその位置に次の歯の歯胚が分化し、歯が欠けている間は両脇の歯が機能します。歯が溶かされ脱落するのは歯の周期性です。歯が溶かされることを歯の吸収といいます。
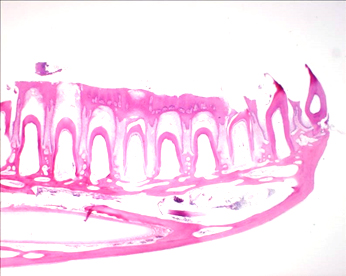 |
 |
イグアナの歯列と歯と顎の結合

イグアナの歯が抜け落ちる時の吸収:歯髄に沢山の破骨細胞がある、しかし脇の歯は何にも変化がない。これは、この歯の周期性によって破骨細胞が出現することを示している。
6 歯は単根ですが側面に溝がある場合が多い。つまり近遠心への歯の分離傾向を示すとも考えられる。また根尖が完全に閉じない、それ故に次世代の歯が歯髄に入るようです。(哺乳類の歯は多咬頭多根が原則;歯の多様性。)
7 様々な形の歯が多いのですが原則として単咬頭(円錐)単根です。
多様な歯の形の一つに牙のような長くなる形をした毒牙があります。
8 歯冠のエナメル質は一般的に両生類よりも厚く(それ故エナメル質形成不全がある)、鋸状縁、皺壁などが見られます。また組織としては、結晶が一定の集団となるcrystal unitや結晶の走向が急激に変わるために哺乳類のエナメル小柱の小柱鞘のような構造も認められます。殆どのエナメル質は無小柱なのですが、大小のエナメル小柱が認められる種類もいます。
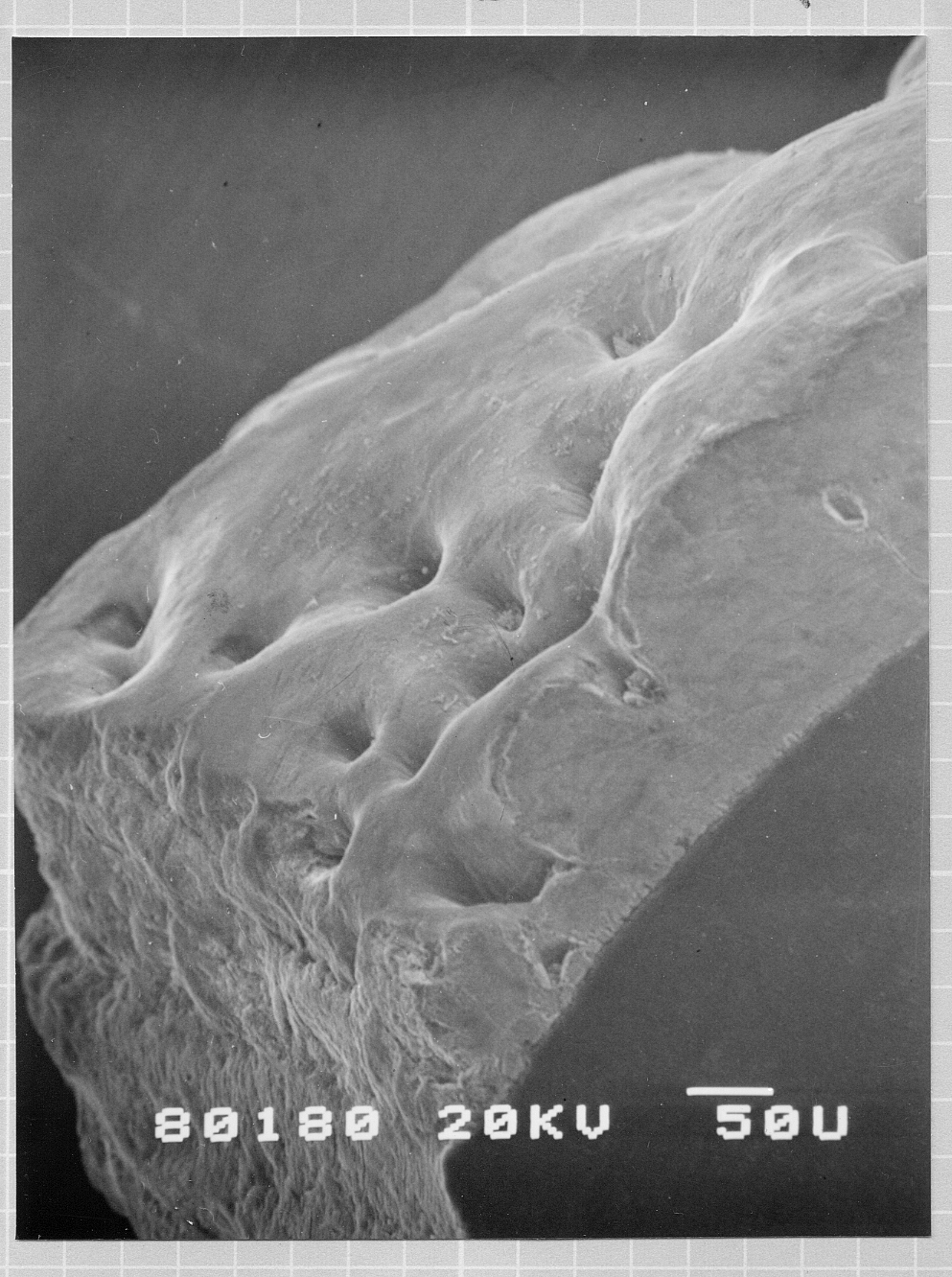 |
エナメル質の形成不全:陥凹の部位が形成不全の部位 |
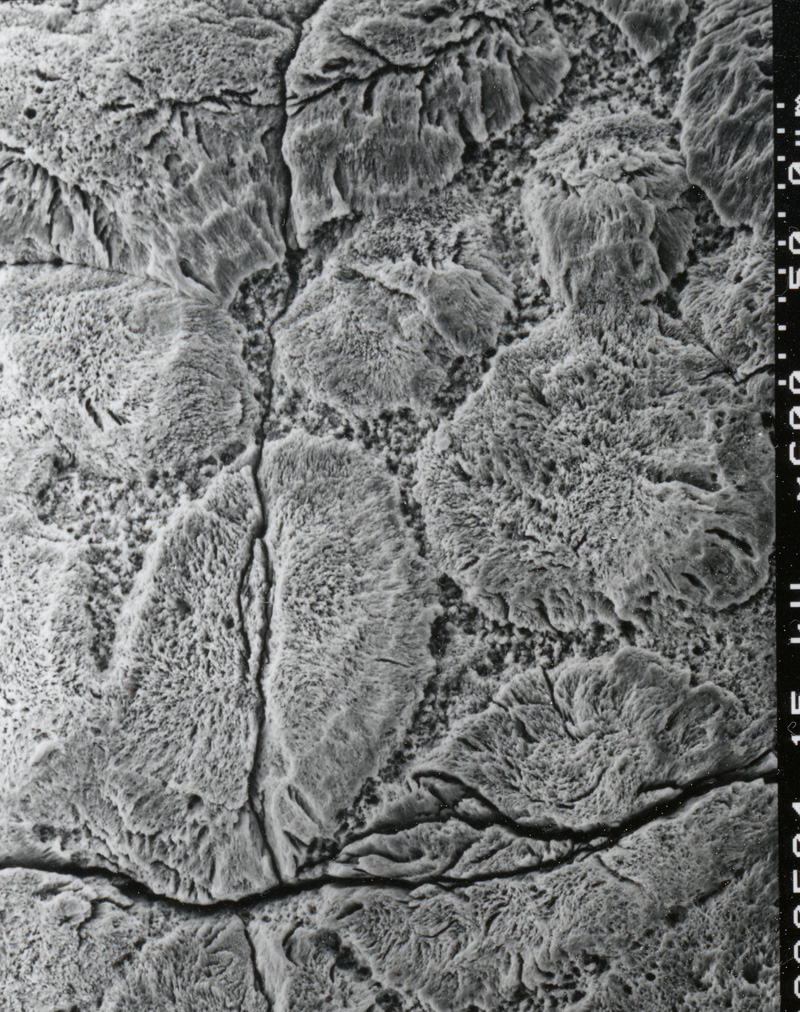 |
 |
ワニのcryatal unit

9 象牙質の象牙芽細胞あるいは象牙細管の集団的模様つまりシュレーゲル条は明瞭ではない。(哺乳類の象牙質ではシュレーゲル条がある。それが発達すると大形の牙の縞模様となる。例;象牙模様。)
10 歯根はほとんどの種類で歯足骨によって顎骨と癒合するが、槽生のワニなどでは歯根膜、細胞セメント質(シャーピーの繊維がある)が発達する。歯根膜は膠原繊維の束(主線維)がありその間に多数のルフィニ小体がある。(哺乳類の歯根はセメント質、歯根膜、歯槽骨という歯周組織によって形成されている。)
歯のない爬虫類
カメ類には歯がなく嘴(くちばし)があります。同様に爬虫類ではありませんが鳥類も嘴がありますが歯はありません。鳥は始祖鳥のみ化石で歯が確認されています。
嘴は口の縁の皮膚が厚く角質化(角化)したもので人間では爪や毛と同じものです。皮膚の細胞の中にケラチンがぎっしり詰まって沢山の細胞が固まって作るものでヤツメウナギの角質歯と同様の構造です(疑問を参照)。ですから基本的に歯と違う器官です。ただケラチンと歯のエナメル質をつくるもととなるアミノ酸の構成は近いものがあります。しかし別の遺伝子によって作られるので別物として考える研究者が多くなっています。議論のあるところです。
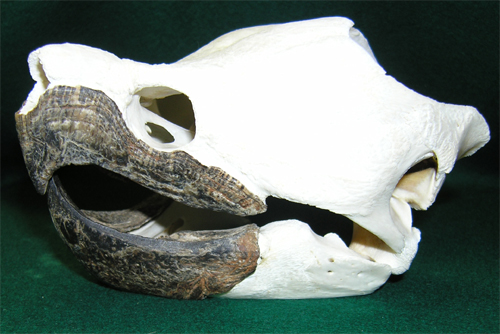 |
ゾウガメの頭蓋:角化した嘴 |