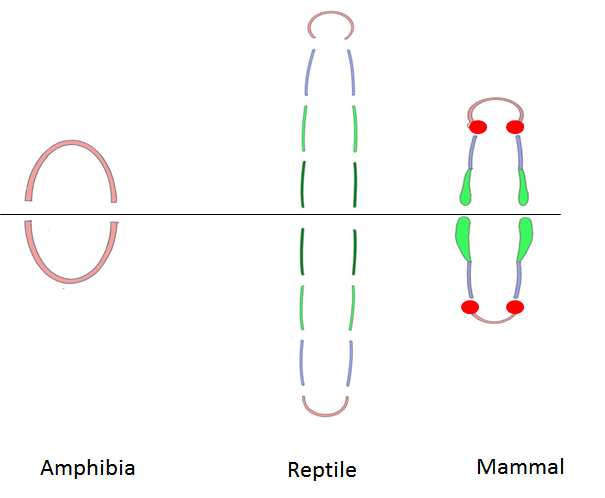
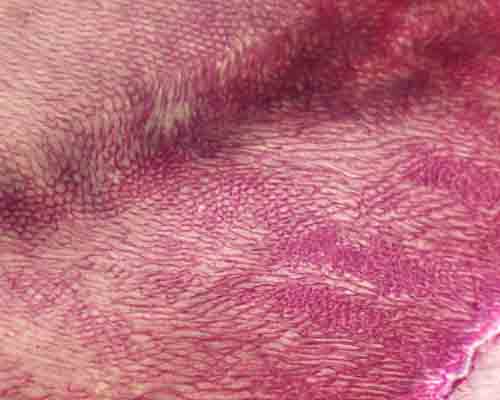
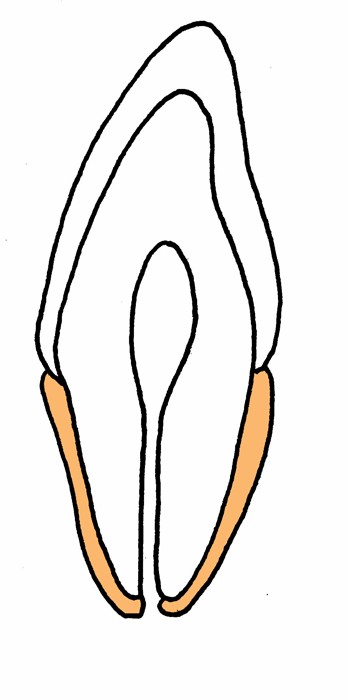
左が両生類で分節が分かりにくい、中央が爬虫類で分節が多い、右が哺乳類で切歯領域(橙色)、犬歯領域(赤)前(小)臼歯領域(紫)、(大)臼歯領域(緑)になることが多い。
目次
1 哺乳類の歯の一般的な特徴
3 歯の形態と形質の特徴
エナメル質、象牙質、歯周組織(セメント質、歯根膜、歯槽骨)
5 上顎骨・下顎骨
6 周期的構造
哺乳類の歯は、哺乳類の種類が魚類や爬虫類と比較して格段に歯数が少ないのにもかかわらず、歯の形が多様で歯種を形成する、体の大きさに比較して非常に大きいこと、そして歯の数と共に交換回数が少ないこと、を特徴とします。ですから、歯数、歯種を区別しやすく種の特徴として歯の形態を利用するということになります。化石の動物は「○○ドン」という名称が良く使われますが、その意味はodonsつまり瘤、突起、「歯」と言う意味からきていますです。
なぜ哺乳類は多様な形の歯を持つように発達したのか?については様々な要因がありますが、一つには我々の目につきやすいために色々な形質が見えることにあります。そして多様な形になる要因の中でも大切なことは外胚葉系の細胞の機能が高度に発達したことが挙げられます。その一つが脳の発達です。脳を動物性器官とすれば、その対称である植物性器官の消化器官の一つである歯の発達ということが対称性の原理(解剖学参照)から理解することができます。
歯冠の表面を覆うエナメル質は皮膚性外胚葉由来ですから質、量ともに多様に発達するのです。これに加えてエナメル質の対称構造の発達、つまり象牙質、歯周組織(セメント質、歯根膜、歯槽骨)の発達を挙げることが出来ます。その結果これらが一体になった構造としての「歯」の多様化が生れるのです。試みにエナメル質を爬虫類のエナメル質と(体と歯の大きさの比率に関係なく)単純に比較してみます。爬虫類は最大でもせいぜい400㎛の厚さです。しかし、哺乳類では、ゾウの先祖のマストドンでは約9-10mm(実に100倍)の厚さに、デスモスティルスは8-10mmの厚さになります。これは歯の大きさから推定して(少なく見積もっても)約数十ないし百万倍以上の容積となります。これを私は脳に次ぐ哺乳類の特徴だと考えています。
歯の大きさにしても同様です。大型のゾウの臼歯は数キログラムもあります(もちろん原始的な哺乳類は米粒ほどの小型の歯を持っているのですから単純な比較はできません)。その上にエナメル質における哺乳類共通の特徴はエナメル小柱と言う組織構造を持つこと、これ以外にセメント質・歯根膜・歯槽骨を持つことを挙げることができます。これは爬虫類では一部に表れる種類もあるのですが、哺乳類では一般的に認められる特徴です。
次に哺乳類の歯の特徴を具体的に観てゆくことにします。
2 哺乳類の顎の分節と歯の数と種類
歯の数を規定するのは一次的には顎の分節であることは爬虫類の項目で書きました、二次的三次的要因には分節相互の関与、分節内の色々な要因が挙げられます。
哺乳類の顎の分節は多くても4分節、あるいはそれ以下となります。そしてそれぞれの分節には特徴を持った歯が形成されるので、これに基づいて各分節を切歯領域(部)、犬歯領域(部)、前(あるいは小)臼歯領域(部)、(大)臼歯領域(部)と呼んでおきます。
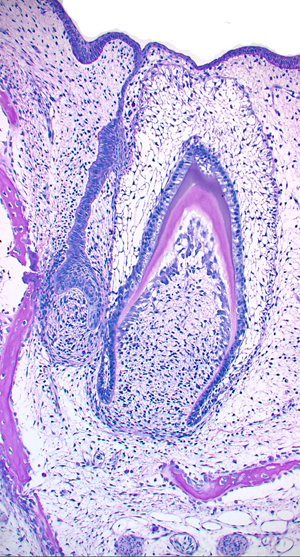
哺乳類では顎の分節は歯堤として現れます。歯堤は、乳歯(乳歯歯堤)で明瞭ですが、永久歯では今ひとつ明らかではありません。歯の発生が一番よく研究されているのはヒトですので、ヒトの発生をいちおう現時点における哺乳類の基準とします。上顎はそれぞれの乳切歯部が2カ所と乳臼歯部、合計3カ所に分かれて生じます。下顎は乳切歯部と臼歯部の2カ所です。ある程度発達すると上顎は乳切歯部が一つとなります。やがて切歯部の歯堤の後端(顎の奥の方、遠心部)で乳臼歯部の歯堤と繋がります。この境界(乳臼歯寄り)のあたりから乳犬歯が分化します。つまり乳犬歯の前後(近心と遠心)2カ所の歯堤から乳切歯と乳臼歯が分化し、それぞれの代生歯堤から永久歯が生じます。つまり乳切歯は永久歯の切歯(代生歯)と交換し、乳犬歯は犬歯と、乳臼歯は前臼歯によって交換します。
しかし、大臼歯のみは乳臼歯部の歯堤の遠心(後)端が顎の生長とともにさらに遠心に伸びて次々と歯胚を分化します。これが大臼歯(あるいは臼歯)です。故に大臼歯は乳歯(と近い関係)であると考えるのが一般的です。
しかし、第一大臼歯と第二大臼歯の歯堤の関係は微妙で、第一大臼歯の後継歯が第二大臼歯、第二大臼歯の後継歯が第三大臼歯であるということも現時点においては否定できないこともここに記しておきます。(これが現実となると哺乳類に三生歯性の証拠となります・・・・・。)
| |
顎の分節の比較 左が両生類で分節が分かりにくい、中央が爬虫類で分節が多い、右が哺乳類で切歯領域(橙色)、犬歯領域(赤)前(小)臼歯領域(紫)、(大)臼歯領域(緑)になることが多い。 |
種による顎の分節の違い
歯堤(顎の分節)の分化部位は哺乳類の種類によって違いが有るようです。これは私が直接研究したゾウやトガリネズミ、ジャコウネズミの歯堤形成から導き出した今の結論です。歯の数や種類、形などは動物の種類によってどの顎の分節を持っているか、またどの分節が発達するのかなどはまちまちです。また上顎と下顎の分節が非対称的な種類も有ります。そして、切歯領域がない種類、犬歯領域が発達する種類、切歯と臼歯領域が発達する種類も有ります。
顎の分節は種類のよって特徴的に発達し、歯の様々な進化と深く関与します。
ヒトの顎の分節は4分節となっていて、各文節ごとの歯が少なくなっています。イヌなどは、切歯領域、犬歯領域、小臼歯領域、大臼歯領域が明瞭に分かれている種類です。一方、ネコなどは小臼歯と大臼歯領域が狭くなっています。ウマなどは各領域が明白なのですが、犬歯領域が広いわりには犬歯の発達が悪く、小臼歯と大臼歯領域が同化して小臼歯が大臼歯化します。ウシの種類は上顎の切歯領域と歯が退化し、歯は歯胚のまま消失しますが、下顎の犬歯(犬歯領域)は切歯領域に入り込んで切歯化します。
ゾウは上顎では切歯領域の近心側は退化するのですが遠心側が犬歯領域の影響を受けて側切歯が犬歯化して象牙になります。下顎の切歯領域と犬歯領域は退化します。臼歯領域は上下顎と共に顎全体に及びます。ラットやマウスでは切歯領域が深くなり切歯が減少しますが常生歯となり、犬歯領域と小臼歯領域は退化して歯も退化するようです。イルカは犬歯領域が顎全体に広まるため犬歯のような歯が顎全体に数十本生えます。またヒゲクジラはイルカと同様なのですが歯が退化します。
 |
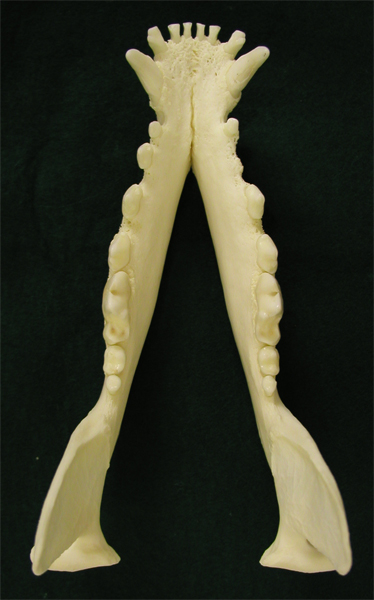 |
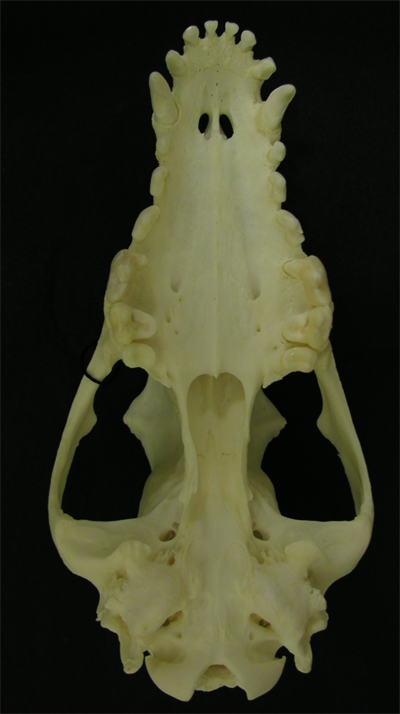
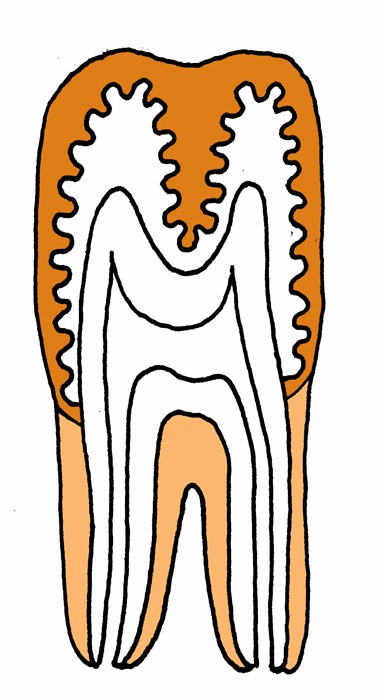
イヌの歯列弓は切歯領域、犬歯領域、前(小)臼歯領域、(大)臼歯領域が比較的はっきりしている。 |
 |
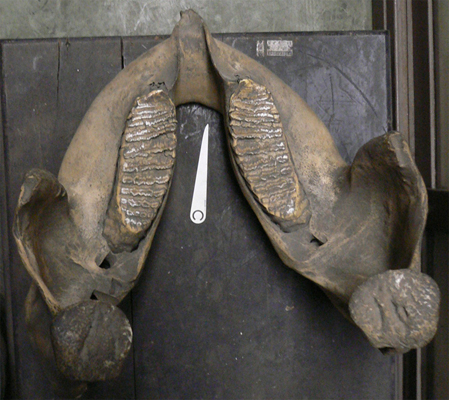 |
インドゾウ(医科歯科大学標本)は上顎は切歯領域に犬歯化の影響を受けた象牙があり、犬歯領域は無く、臼歯領域が顎の大半を占める。下顎は切歯と犬歯領域が無く、臼歯領域が顎のほとんどを占める。前(小)臼歯領域は(大)臼歯領域と癒合する、と考えられる。 |
歯の減少の要因
哺乳類の歯の数が爬虫類などより何故減少するのかについて、顎の分節の減少によることは有る程度分かったと思います。では、何故顎の分節が減少するのでしょうか?これは体全体を観ないと解りません。化石などと現在生きている動物を比較すると、体の数の骨の減少を挙げることが出来ます。つまり体全体の分節が減少し、その代り一つ一つの分節が複雑になっている、と言うことです。それは体の器官が高度に複雑に発達することと関連しています。
哺乳類に限らず生物(生命体)は、つねに体が発展(進化)するという大原則があります。哺乳類における発展(進化)の一つが脳や歯の複雑な進化ですが、それを保証する現象(対称によるバランスの維持)が分節の減少、と考えることが出来ます。体の分節の減少と関連して歯に起こった現象が歯の数の減少なのです。むろん歯には歯それぞれの減少様式はありますが、顎の分節の減少深く関与している、と言うことです。
歯の数の減少は、顎の分節の減少と関与しますが、どこの分節が退縮するのかは種の特徴です。そして分節の中の歯の数もまた減少します。歯数の減少を対称的に補う(保証する)のが多様な歯の形態分化です。多様な顎の分節や歯の形態分化は分節や歯の減少によって補(保証さ)れているとも言うことができます。
3 歯の形態と形質の特徴
哺乳類の歯の形態は様々な形となります。換言すれば歯は様々な形質によって作られるという特徴があるとも言えます。
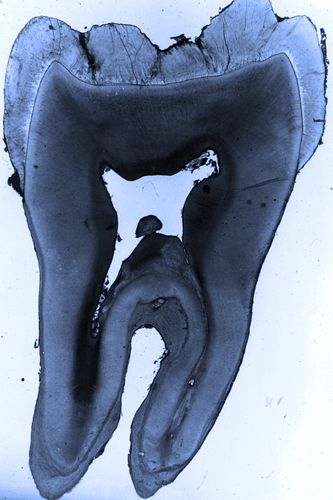
まずエナメル質が厚く発達するので歯冠と歯根がハッキリと区別出来ます。
歯冠の表面には様々な形の膨らみがあります。一般的には大きな独立した膨らみを咬頭と呼びます。膨らみが山の尾根あるいは堤防のような形となるときは隆線(あるいは堤)と呼びます。小さな膨らみを結節(あるいは副咬頭)などと言います。歯冠の縁の高まりを辺縁隆線と呼びます。しかし歯は動物の種類によって形が多様なためこの用語には決められた規則はありません。此処に記したのはヒトを中心とする用語で、ヒトの歯が詳しく研究されているため、比較的使いやすいという面があります。しかしそれぞれの動物によって使い方にそれぞれ習慣がありいりいろいろな名称を使い分けています。
注:ほとんどの研究者は、咬頭や結節を歯の進化の仮説から命名した用語法を用いています。しかし、私は(後で述べますが)歯の進化の考え方に対して違った視線をもっていることもありますが、より誰にも分かりやすい方法(歯科医が使っている名称)を用いています。これによって殆どの哺乳類の歯の形質が同じ土俵の上で議論し理解できます。
歯根は複数に分岐することが多く、歯根の表面には溝や窪み(陥凹)が出来ます。しかしこれらには決まった名称がありません。そこで私は歯冠と同様の方法(歯の中の位置で呼ぶ)で名称を付けて使っています。
咬頭と歯根の定義
歯冠と歯根は対称的な構造ですが、歯冠と歯根を形成する咬頭と各歯根を分析すると、歯根には咬頭が、咬頭には歯根が必ず対応して発生します(下記)。もっとも発生の途中で消失したり癒合したり、または分離することはあります。しかし、それでも発生の初期には咬頭と歯根は対称的に分化します。もちろん比較解剖学的にも大きな咬頭と呼べる隆起には歯根が伴います。つまり、系統発生でも歯の発生でも同様に、咬頭へと発達すると歯根が分化し、逆に歯根が発達すると咬頭が伴うという対称的な分化が認められます。ここから「咬頭と歯根は対称的な構造である」という定義が生まれます。つまり「咬頭は歯根を伴い、歯根は咬頭を伴う」、と言うことです。これは、咬頭と歯根がより複雑化した哺乳類の歯の明瞭な特徴といえるものです。
 |
マンモスの臼歯 歯冠(下側)の表面はエナメル質の上を歯冠セメント質が覆っている。亀裂はセメント質のもの。上は歯根で表面に横方向に走るスジ(根周条)がある。歯根は歯冠の楯の筋(咬頭に近い構造)に対応している。左端では複数の咬頭に対応する歯根が癒合して一本となる。(ここで言う咬頭はゾウでは咬板と呼ばれ複数の咬頭が癒合したものと考えられている。) |
歯髄腔と歯髄
歯の中心には歯髄腔という空間があり歯髄が入っています。歯髄は象牙質やエナメル質、セメント質とは違い細胞や神経、血管が入っています。歯髄腔の形も特徴があるのですが、この形に注目した研究が少なくあまり解明されていません。ただ一般的にいえることは、ヒトを例にとると、切縁(切歯)、尖頭(犬歯)、咬頭に対応して髄角がある、咬頭に対応する歯根には根管がある、とい言うことです。そして大切なことは、歯髄腔が歯冠(歯冠歯髄)と歯根(歯根歯髄)とに、そして歯根歯髄は、根幹、歯根中腹(中央部)、根尖部とに3区分されると言うことです。さらに根尖部の先の根尖端は曲がったり細くなったりと非常に変化に富んだ部位ですが、これを区分することもできます。この区分は歯根の外形と連動します。
以上をまとめると、咬頭と歯根の関係と同様のことが歯の外形と歯髄にも認められると言うことです。
歯冠と歯根の対称性の発生学的根拠
歯の原基(歯胚)の発生において咬頭に対応して歯根が分化します。これを咬頭と歯根の対称性、そして、体の対称性(体制)の視点から考察すると、歯冠が体の表面へ(方向)分化し、これとのバランスつまり安定性から、歯根が体の深部(方向)へ分化する、と言うことになるということは上で述べました。これが哺乳類の歯の発生でハッキリと現れるのです。すなわちエナメル質を形成するエナメル器は表面に向って分化し厚いエナメル質を形成します。それと対称的に歯根を形成するヘルトヴィッヒの上皮鞘は深部へ向かって分化し、しっかりとした歯根を形成します。
この現象は、系統発生的に、そして体制的には次のように理解されます。サメを除いてほぼすべての歯には明瞭な歯冠と歯根があります。魚類の歯の尖端は象牙質と同じ由来を持つエナメロイドで形成されているとはいえ、エナメロイドは上皮の強い影響によって分化した構造であり、かつ形態的にも歯冠と言うことができます。これに対して象牙質は、エナメル質(エナメル器)に接して形成され、皮膚の深部へ分化して歯根をつくります(魚類の歯参照)。象牙質はエナメル質と接して分化する構造であり、皮膚では表皮(エナメル質)に接する真皮に例えることができます。それゆえエナメル質は上皮に由来し、象牙質は真皮に由来すると、理解することができます(これは様々な組織学的な特徴からも証明できます)。
そして歯根の本態は表面にエナメル質を伴わない(セメント質で覆われた)象牙質ですから、歯根は真皮に由来する皮膚の深部へ向かって分化した構造と言うことになります。エナメル質と象牙質はそれぞれが別の方向へ向かって分化する対称構造なのです。つまり歯は基本的に皮膚の構造と対応する、皮膚の構造が変化した器官ということです。それゆえ、エナメル質と象牙質つまり咬頭と歯根は系統発生学的にも対称的構造であるわけです。
以上の事実は歯冠と歯根は対称的構造が体(体制)の構造と同じ法則で歯も形成されている、と言うことを示しています。
注:これは哺乳類の歯を観察するときに歯冠と歯根、そして歯髄の形態を関連付けながら行わなければならないこと、さらに顎の形態と歯の形態も同様に観察する必要があることを示しています。しかし、現在の歯に関する本とくに教科書ではこのような関連において記述しているのは殆どありません。
 |
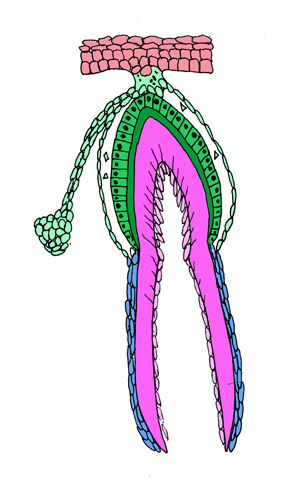 |
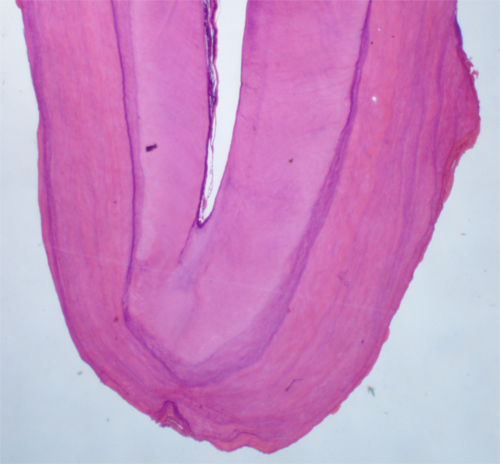
ここでは理解しやすいようにワニの歯の分化で説明する。 左の写真はワニの石灰化した歯胚(右)とその次の世代の歯胚(左)。歯胚から上に伸びるのが歯堤で上にある口腔粘膜上皮につながっている。これを右図でしめすと、橙色が口腔上皮、黄緑が歯堤と歯胚、こい緑がエナメル質とエナメル芽細胞、青色がヘルトヴィッヒの上皮鞘、濃いピンクが象牙質となる。 エナメル質は口腔上皮の歯胚から分化したエナメル芽細胞によって作られ口腔粘膜(上側)へ作られる、象牙質(歯根象牙質は ヘルトヴィッヒの上皮鞘に誘導された象牙芽細胞(薄いピンク)によって粘膜下組織(下側)へ形成される。歯冠と歯根の対称的分化を示す。 |
4 歯の組織
エナメル質
哺乳類のエナメル質は厚く発達するとともに、構造が複雑になります。まず一般的に認められるのはエナメル小柱という結晶の集団です。これが象牙質との境からほぼエナメル質の表面まで続きます。エナメル質には細い管があります、これをエナメル細管と言います。原始的な哺乳類ではエナメル小柱と同様に象牙質との境目からエナメル質表面まで続きます。しかし多くの種類では象牙質表面にだけ存在するため、エナメル紡錘あるいは棍棒または(単純)突起などと呼ばれています。
 |
 |
 |
 |
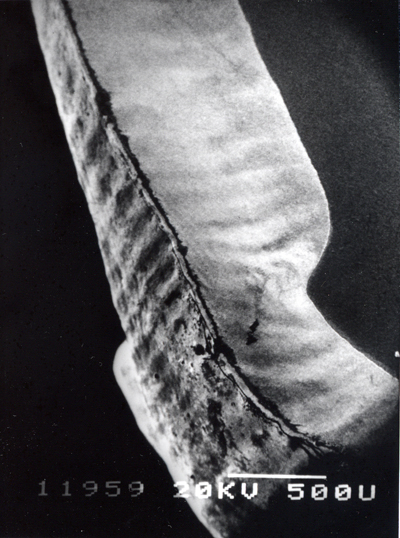 |
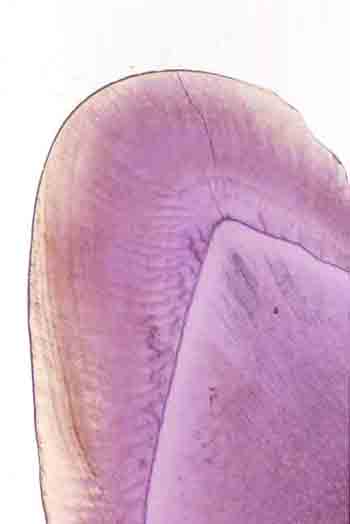
歯の全景(左上)、エナメル質の咬頭の部分(中央上)には横に湾曲して走るハンター・シュレーゲル条が観察される。その一部の拡大(右上、その右の下に一寸象牙質が覗いている)は米粒のようなエナメル小柱がこの構造を作っていることが分かる。咬頭の下のエナメル質と象牙質の境界(左下)には象牙細管(下側)からエナメル質(上側)に入っているエナメル紡錘が沢山あることが分かる。 |
エナメル小柱はまっすぐに配列する種類もありますが、ほとんどの哺乳類では象牙質との境界からエナメル質表面に向って曲がります。しかもエナメル小柱は集団にまとまることが多く、この集団も曲がる、つまり「うねる」ようにしてエナメル質の表面まで配列します。それ故に様々な模様がエナメル質の断面に現れます。これをハンター・シュレーゲルの条紋(あるいは単にシュレーゲル(の)条(紋))と呼びます。
エナメル質は周期的に形成されるため年輪のような成長線が出来ます。この成長線はエナメル質の表面に周波条というスジを作ります。
このようなエナメル質を構成するのは殆どハイロロキシアパタイトの結晶ですが(歯学参照)、それ以外にエナメルタンパクや水分などが含まれています。
象牙質
脊椎動物の象牙質はサメなどと基本的に同様の構造をしています。象牙細管が歯髄腔の表面からエナメル質との境界まで続き、その細管には歯髄に位置する象牙芽細胞の突起がはいり、ときにはエナメル質まで侵入し、象牙細管がエナメル細管と連続します。
さて、哺乳類における象牙質の特徴は、エナメル質と同じように発達して象牙やセイウチの牙のように巨大な歯に発達するための基本的な成因となることです。この基本的な成因が、象牙細管によって分かります。これは象牙細管が曲がりくねってエナメル質表面まで達することと、さらに象牙細管が集団になり全体として「うねる」様な集団的曲走(つまりうねり)をすることです。この時、様々に変化した象牙細管が模様を作り出します。
さて直接に歯の巨大化を支えるのは象牙質そのものの構造(基質)です。これは象牙質の基本的な構成成分である膠原繊維と結晶の建築学的構造を強度に依存します。この基質の構造が象牙芽細胞の集合と動き(ダンシング)によって作られるのです。つまり、象牙細管のうねりなどの変化に沿って膠原繊維や結晶が配列します。このようにして象牙細管の模様にあらわされるような基質の構造が形成され、象牙質の強度が強くなり巨大化する歯が保たれているのです。
象牙質のこれらの構造(模様)はエナメル質同様にシュレーゲル(の)条(紋)と呼ばれます。シュレーゲル条は象牙質の断面では美しい模様となるため、「縞」(striation)あるいは象牙で特徴的なので象牙模様などと言われます。象牙細管もまた規則性をもち周期的に作られるためこれらの構造は比較的規則的な模様となります。この周期性は大きな歯ほど認めやすく小さな歯ほど認めにくいので、ラットやマウスを用いることが主流となっている今の研究では余り問題にされていないという現実があります。それ故あまり教科書には書かれていませんし、また解明も進んでいません。
周期的に象牙質が作られることは象牙質にもエナメル質同様に成長線が形成されることです。様々な成長線が観察されるため、いろいろな成長線の名称がありますが、基本的なのは年輪と同様に同心円状となるもので、アンドレーゼン線、オーウェン線、象牙層板などと呼ばれています。そのうち象牙質表面にスジ模様となるのを根周条といいます。基本的にはエナメル質の周波条と同様の機構によって形成される構造です。
注 象牙質の成長線の名称は観察方法によって見え方が違うためです。
 |
 |
左は井尻先生の象牙のパイプ、右はカバの犬歯の断面 共に縞構造がある。 |
 |
縞構造の原因は象牙質にある象牙細管の周期的な曲走と配列、太さの変化、其処から出る側枝の角度の違いなどです。この変化と膠原繊維、結晶の配列が対応して全体的に縞模様が出現します。(シュモール染色) |
歯髄
歯髄は歯髄腔に入っている細胞などの名称です。象牙質との表面には象牙芽細胞が一列に配列して、その突起が象牙線維として象牙質の細管(象牙細管)に入っています。象牙質と象牙芽細胞の境界にはまだ石灰化していない軟骨状の象牙前質があります。象牙芽細胞の内側(歯髄の中央側)には歯髄細胞(繊維芽細胞)が沢山ありますが、ウニか星のように突起をたくさん出して突起同士が手を結んでいます。ですからスポンジのような状態です。スポンジの隙間には膠原繊維がまばらに詰まっています。
歯髄には根尖孔から入ってきた神経と血管もあります。この分布は歯種、動物の種類によって違います。
歯周組織
歯を顎と結びつけかつ咀嚼の圧力の干渉となるのが歯周組織です。哺乳類では歯根の表面からセメント質、歯根膜、歯槽骨、歯肉とつづきます。歯肉は口を開けたとき歯の周囲に見える歯茎のことです。これらの組織は歯とともに発生しますが、これらの土台となるのが顎の骨です。ヒトの場合、上顎では上顎骨、下顎は下顎骨です。ちなみに歯槽骨は顎の骨と骨癒合をしますが、上顎では歯槽突起、下顎では歯槽部という別々の名前が与えられています。
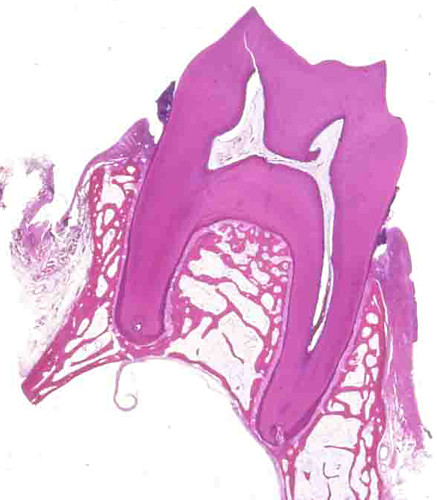 |
上顎の臼歯の断面(脱灰標本の為エナメル質はない) 上顎のため実際は天地逆で下の空隙が上顎洞となる。歯の中心が歯髄、歯根の周囲にある骨(歯槽骨)との境に歯根膜がある。歯根の表面にはセメント質がやや濃い紫色の層となっている。歯槽骨の表面には歯肉がある。 |
セメント質
セメント質は基本的に歯根周囲を覆う骨に似た組織です。骨との最大の違いは、骨は常に壊され(吸収という)また作り直される(再生という)ことを繰り返しています。これを改造現象あるいは単に改造と言います。しかしセメント質は改造がありません。セメント質は加齢によってどんどん厚みを増します。ある種の哺乳類ではセメント質はエナメル質の表面まで覆い被さるように発達します。これを歯冠セメント質と言います。セメント質と骨の共通点は、基本的に膠原繊維の間にハイドロキシアパタイト結晶が
沈着し、無機基質(アパタイトの結晶がほとんど)が約60%であること(象牙質より少ない)、そして細胞(これをセメント細胞と骨細胞といいます)が骨の中に並んでいることです。顕微鏡で骨を見ると骨細胞が、様々な形と大きさをしていますがほぼ同心円状をなして配列しています。これを骨単位(骨層板)と言います、しかしセメント質にはこれはありません。セメント細胞は曲線状に配列することはあってもほぼ一列になります。ただ歯冠セメント質では骨単位のような構造(セメント単位)をたまに認めることがあります。しかし、骨単位とセメント単位では作られ方が全く違います。骨単位は改造現象のため外側から内側へ作られる(吸収した空間に骨が作られるために、壁側から骨が再形成される)のにたいして、セメント単位はセメント質が(多くの場合は血管などを中心にして)同心円状に外に向かって作られるためです。セメント質には基本的に改造がありません。
では、骨とセメント質のこのような違いは何なの?、でしょう。それは体の中の形成部位の違いと私は考えています。セメント質は象牙質、あるいは歯冠をつくるエナメル質を覆うようにして形成されます。この位置は体の構造からいえば、真皮の下層、皮下組織との境界に当たります。つまり組織学では真皮の網状層と言うことになります。ここに形成される硬組織がセメント質なのです。網状層は膠原繊維、平滑筋繊維、弾性線維が豊富にあり密生結合組織(強靭性結合組織)に分類されています。ここでは骨が形成されてもそれほど大きく分化することが出来ません。硬組織は、表皮に近いほど表皮の影響を受け、遠いほど影響が少なくなります。ですから皮下組織に出来る骨と真皮に出来る象牙質の違いが生れ、その間には移行的なセメント質や歯足骨が形成される、と言うことになります。ですから歯足骨に弾性繊維があるのも納得できるという訳です。
 |
 |
左が歯根の表面を覆うセメント質(橙色)の模式図、右が歯根の先端を覆うセメント質。(加齢によってセメント質は厚くなる)、 |
 |
 |
左が歯根セメント質(薄い橙色)と歯冠セメント質(濃い橙色)の模式図、右がゾウの臼歯エナメル質(下)の表面に形成された歯冠セメント質。 歯冠セメント質は塊状に形成される(沈着する)こともある。 |
歯根膜
歯根膜は歯根の周囲にあり歯周靭帯とも呼ばれる膠原繊維が主体となる組織です。膠原繊維はセメント質と歯槽骨の中に埋め込まれ、この間にハンモック状に張っています。膠原繊維によって咬む圧力や歯を引っ張るちから歯を保持します。歯に対する圧力や引っ張る力を感じるセンサー(感覚受容器のルフィニ小体)が膠原繊維と密接な関係を持って位置しています。痛みを感じる自由神経小体も豊富にあります。そして血管が歯根を取り囲むように走行配列し、おおくが神経と伴行します。
歯根膜は歯が破損して抜けたとき(脱臼といいます)、再び元の位置に戻すと石灰化してセメント質と歯槽骨が骨性癒合してしまいます。しかし、通常ではなぜ石灰化しないのか、と言う疑問はまだ解決していません。比較解剖学的に言えることは、両生類や爬虫類の歯足骨周囲の線維性組織が歯の形態進化と相応して歯根を取り囲むようになった、と言うことです。歯根にある神経、血管も同様にして変化したことが分かっています。
 |
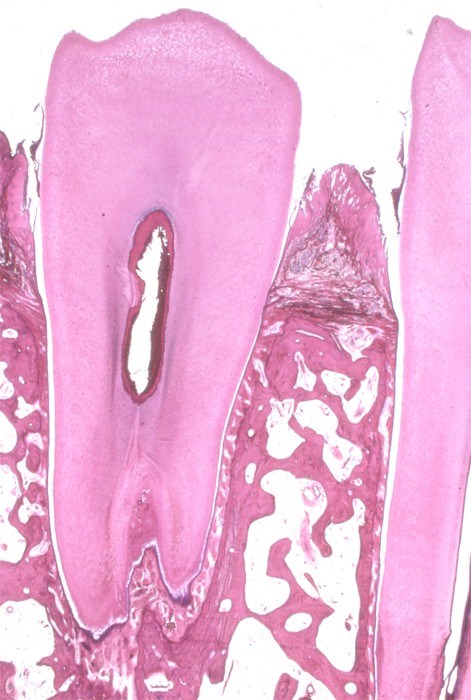 |
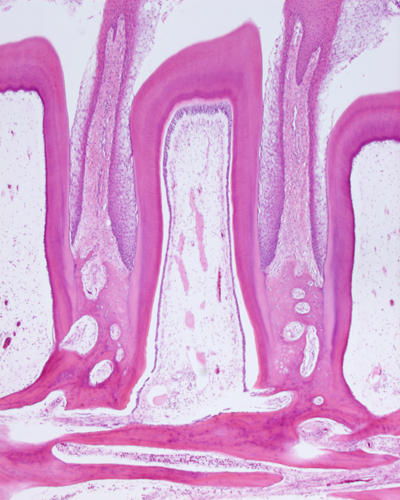
左はイグアナの歯根と周囲組織、右はヒトの歯根と歯周組織 イグアナの下方の顎骨と歯根と歯根を結ぶ歯足骨は明らかに構造が違う、しかしヒトの顎骨と歯槽骨はそれほど違いがない、しかしセメント質、歯根膜、歯槽骨の歯根膜面の固有歯槽骨の存在が哺乳類では特徴的です。 |
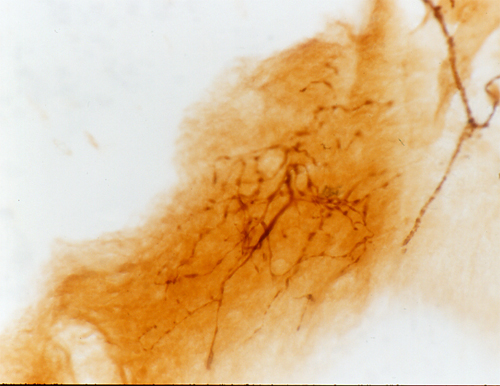 |
根尖の先端にあるルッフィニ小体と言われる神経終末 |
歯槽骨
歯槽骨は殆ど骨と同様の組織ですが、歯根膜(歯根)側の一層は歯根膜から入ってくる膠原繊維を含んでいる層(骨の中にある膠原繊維をシャーピーの繊維という)を繊維骨(あるいは篩板、歯槽硬線、固有歯槽骨)とよび、その外側(歯肉側)のシャーピーの繊維の含まない層を支持歯槽骨といいます。繊維骨は骨単位ができない(セメント質的な)改造が行われます。つまり歯根膜の表面側が吸収や再形成します。それゆえ吸収された跡(ハウシップ窩)が成長線として残りますこれを反転線と言います。骨単位ができないため殆ど平行に(彎曲することはありますが)成長線が走り、これを横切るようにシャーピーの繊維が走ります。これにたいして支持歯槽骨はほぼ骨組織と同様で骨単位、骨髄などが有ります。ただ歯肉に接する面は平行に成長線が走る緻密質骨の層となります。固有歯槽骨はセメント質に似ていますが、支持歯槽骨は皮下組織に形成される骨組織に類似するのです。このことからも歯の構成要素が皮膚の構造と関連性を持つことが分かります。
5 上顎骨・下顎骨
哺乳類の顎骨は、鰓弓(顎骨弓)として上顎は切歯骨と上顎骨、下顎は歯骨(下顎骨)だけで構成されます。爬虫類とそれより原始的な動物の顎骨弓の構成要素がそれぞれの顎の骨に含まれているという議論もありますが、ヒトや高等霊長類では切歯骨と上顎骨も発生の過程で癒合をして一つの骨(上顎骨)となります。この骨の中でそれぞれの歯に分布する神経や血管が分岐して歯の周囲に達すると網目状に分布します。
筋は下顎骨に停止する咀嚼に関する筋だけがあります。上顎骨に関する筋は有りません。ただし咀嚼は顔の表面に位置する表情筋も関与して行われます。下顎に関する筋は、深頭筋群(咀嚼筋群)という顎を上顎の方向へ引きつけ前後左右に動かす筋、下顎を引き下げる(口を開く)ことに働く筋(舌骨上筋群、舌骨下筋群:下顎を開くのは舌骨が支点となる)が有ります。歯を使う咀嚼はこれ以外の筋も共同して働きます。例えば舌(内舌筋、外舌筋)、口唇(口輪筋)、頬(頬筋)などです。そして嚥下には、軟口蓋を構成する筋(口蓋垂筋、口蓋帆張筋、口蓋帆挙筋、咽頭筋など)です。
哺乳類では歯を使う捕食、咀嚼、嚥下は一体となって機能します。ですから、顔全体の表情を鍛えると咀嚼嚥下機能が増して、おいしく食べられると言うことになります。
6 周期的構造
哺乳類の歯はここに示したように様々な面で多様性に発達します。それ故様々な構造がありますが、それぞれの構造を作り出す大切な要因の一つが周期性です。周期性は一般的には成長線として歯をはじめとした器官に印刻されるために様々な成長線の名前が与えられています。しかし周期性は構造自体を作り出すと言う面も忘れてはなりません。そのいくつかを列記します。
エナメル質 日周期(概日周期)の横紋、週あるいはそれ以上の周期の平行(褐色)条(レッチウスの線条)、レッチウスの線条に対応するエナメル質表面の周波条、出産時の成長線(レッチウスの線状の一つ)の新産線(新生線)などがエナメル質の断面や表面に観察されます。これらは基本的にエナメル質を構成するエナメル小柱の石灰化の善し悪しがエナメル質全体としての成長線となるのです。
そしてエナメル小柱は石灰化の程度が周期的に変わるだけではなく、太さを変え、捩れ、波打つ(彎曲する)という変化を周期的に示します。そのうえエナメル小柱は集団となり、エナメル小柱の集団も捩れ、彎曲し、集団の大きさも癒合し分離します。これらもまた周期性を持って作られた構造なのです。これがエナメル質のシュレーゲル条なのです。
象牙質 象牙質は周期的に石灰化の程度が変わるために様々な成長線の名前が与えられています。しかし、象牙の部位によって観察される成長線が違うためにそれぞれの名前は象牙のどの位置にあるのかということがいつも問題になります。一日の周期(概日周期)と言われる石灰化条(同心円のようになることもありリーゼガング現象だという意見もあります)、週あるいはそれ以上の周期のアンドレーゼン線(歯冠象牙質)、アンドレーゼン線の太くてはっきりした成長線のオーウェン線(あるいはオーウェンの外形線、歯冠象牙質)、象牙層板(歯冠象牙質と歯根象牙質)、歯根表面の成長線の根周条などです。
エナメル小柱と同様に象牙細管も太さを変え、捩れそして彎曲し、側枝の派出方向や太さ数を変えます。象牙細管からは側枝という枝を多数出しますが、この側枝の出す数、角度、つまり分岐の状態が周期的に変化します。この変化は、象牙質の膠原繊維の配列、結晶の沈着と関係有るらしく石灰化の善し悪しに連動して上に上げた様々な成長線となるようです。そして象牙細管は集団となり、集団もまた捩れ彎曲します。象牙細管集団の捻れや彎曲はシュレーゲル条(縞模様)となります。これらの成長線やシュレーゲル条が歯根象牙質表面に現れたのが根周条だと考えられます。
<石灰化条>
<化石の石灰化条>
<縞>
セメント質・歯槽骨 セメント質も歯槽骨も骨と同様の成長線があります。これらは一定の周期によって成長線が形成されるとともに細胞が埋め込まれた構造です(埋め込まれた細胞をセメント細胞あるいは骨細胞という)。これを層板あるいはセメント層板、骨層板といいます。そのなかで太い成長線を休止線(骨の成長が一時止まると言う意味)、ハウシップ窩の跡を反転線と言います。支持歯槽骨や顎骨の成長線は比較的平行になっているのですが、とくにヒトのセメント質や固有歯槽骨の成長線は成長線相互の間隔が不規則です(性周期の一定な動物では成長線の間隔が規則的なため、これの原因は性周期と関係すると言われています)。