4 系統発生と個体発生の関係
「系統発生は進化であり地球の歴史とおなじ歴史的時間の現象、個体発生は現在時間の生物の現象である」
「地球規模の原則、法則にのっとった個体発生と系統発は互いに反映する
系統発生と個体発生の関係は進化を論じ理解するために欠かすことができない命題です。
それは、現在は過去の鍵(過去の歴史)であり、過去は現在の鍵(現在)である、からです。これを比喩的にいうなら、何らかの病に冒されたとき、その人の体質(現在)とともに過去や家族の病歴(歴史)をしらべ、それを将来の健康の参考にする、ということです。
E.Haeckel(1866)が、系統発生と個体発生の関係を(巷間にいう)「反復説」として提唱して以来、この学説がさまざまなかたちで生物学的あるいは古生物学的の現象についての説明基準となっているのですが、両者の関係についての議論が侃々諤々と続いています。これは、系統発生と個体発生の関係が成り立つ根拠を立証した研究がないからです。私の見るところ、両者の関係は曖昧のままで、混乱していると感じられるので整理して見たいと思います。
その原因の第一点は、まえに記述した時間の捉え方の錯覚です。つまりなんとなく個体発生は万年の単位で繰り返される、という期待感です。第二点は、それゆえに、立ち位置の違い(例えば古生物か発生学かなど)により、当然と言えば当然なことですが、さまざまな現象、とくに発生現象において、系統発生が反映している、あるいはしていないという見方の違いが生じているのではないか、というものです。
結論からいえば、進化を論じるうえでは個体発生と系統発生は明瞭に区別し、そのうえで両者の関係を整理し、統一することが必要です。そのさいどのような根拠で時間軸の違う両者が関連性を持つのかを明確にすることが大切だ、ということです。
系統発生と個体発生の関係に関する二大学説
両者の関係はさまざまな解説あるいは主張がありますが、私は、Haeckel(1966)の最初の命題と井尻正二による提唱がもっとも本質を突いており、大切なものだと考えています。それゆえこの二つを見てゆくことにします。
まずE.Haeckel(1866か)から見てゆきましょう。彼は「個体発生は系統発生の短縮された、かつ急速な反復であり、その反復は遺伝(繁殖)および適応(栄養)の生理学的な機能によって規定される」としました。いわゆる「反復説」です。
注 http://ja.wikipedia.org/wiki/によるE.Haeckelの訳
「個体発生
(ontogenesis) 、すなわち各個体がそれぞれの生存の期間を通じて経過する一連の形態変化としての個体の発生は、系統発生 (phylogenesis) 、すなわちそれが属する系統の発生により直接規定されている。個体発生は系統発生の短縮された、かつ急速な反復であり、この反復は遺伝および適応の生理的機能により条件付けられている。生物個体は、個体発生の急速かつ短縮された経過の間に、先祖が古生物的発生の緩やかな長い経過の間に遺伝および適応の法則に従って経過した重要な形態変化を繰り返す」
私達は、生物の進化をほぼ信じているうえ、さらに動物園などでもサルとヒトの子供の共通性をみているためこの反復説は非常に判り易く腑に落ちるものとなっています。
歴史的にみれば革命的な「種の起源」がかのダーウィンによって著され、その論拠ともなる個体発生と系統発生の関係がもたらされたのです。種の起源による進化論と反復説は当時の学会、むろん色々反論はあるにしても、今日に至るまで世界を席巻していると私はみています(現在もこの強い影響下にあります)。
ですから結論的に言えば、それ故にHaeckelの考えは、「そのとおり」と錯覚しやすいもの、ではないでしょうか。この錯覚はあとをよむと分かります。
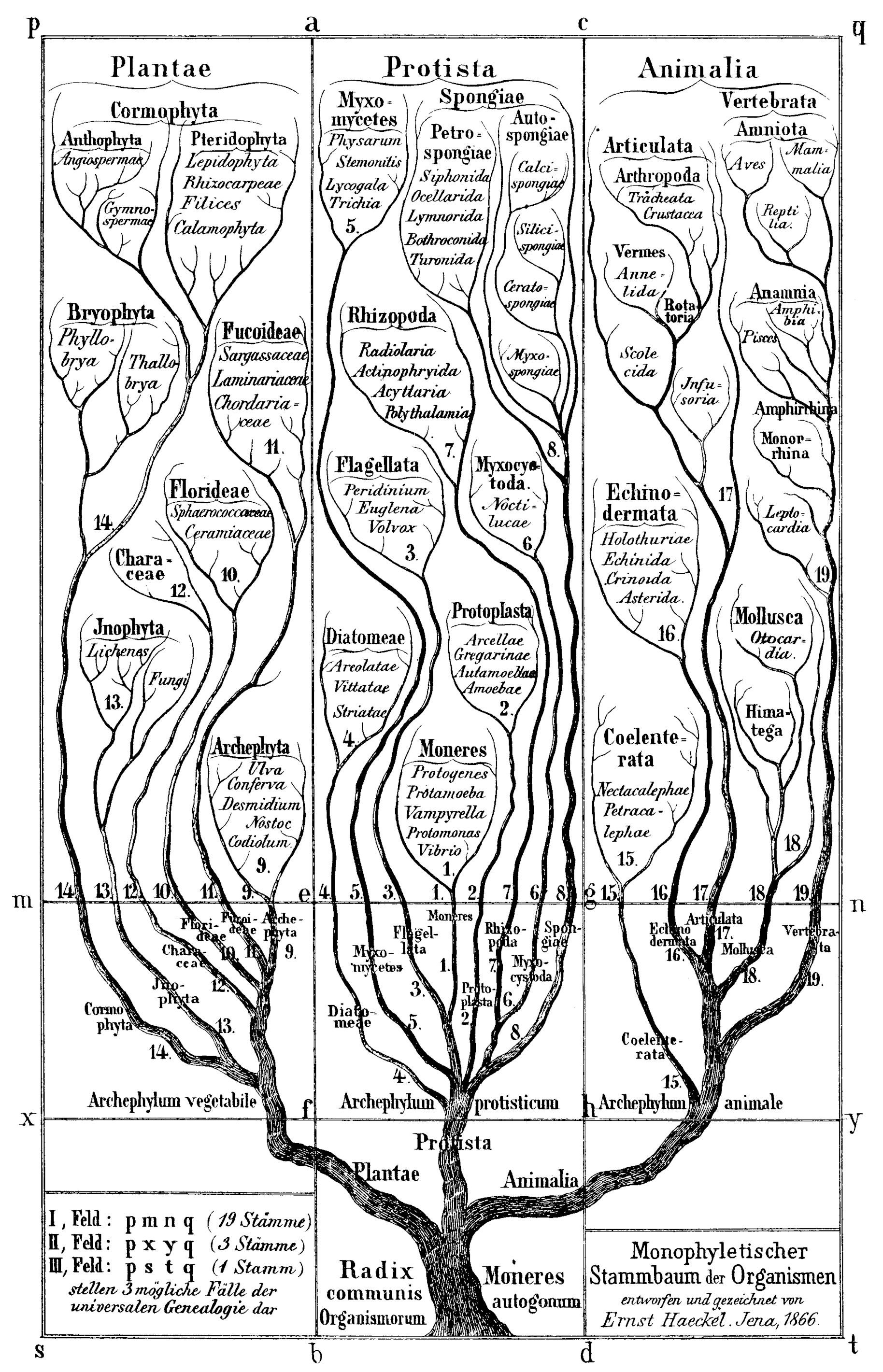
ここで忘れてはならないのは、Haeckelが膨大な生物の多様性、さまざまな発生、化石を研究したことです。そのうえで系統樹をつくりこの大命題を提唱したのです。第二点は彼がこの命題を打ち立てたときは、かのC.Darwinが「種の起源」(1859)を著し、大方の科学者はこれに圧倒されこの進化論で説明しようとした時代、種の起源の大津波が押し寄せた時代だった、ということです。
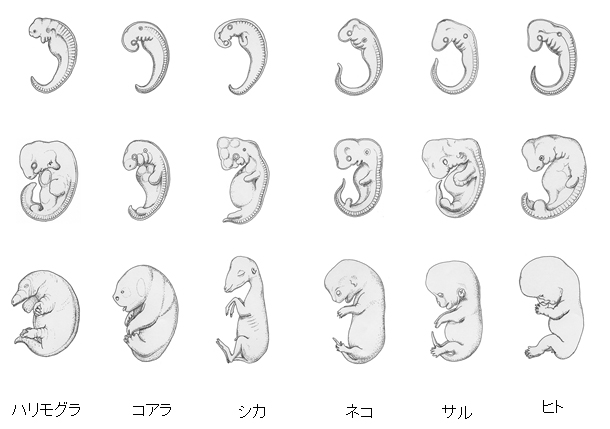
ヘッケルの胎児の発生形態の比較と系統図
|
|
 |
一方、井尻(新版 科学論(下)、38頁、大月書店、1977年)は「個体発生は系統発生によってあたえられ、系統発生は個体発生を通じて展開する。」つまり、「個体発生の合法則性(論理性)は、系統発生の必然性(歴史性)によってあたえられ(獲得され、刷り込まれ)、系統発生の合法則性(論理性(=必然性)は、個体発生の偶然性(変異性)を通じて展開(生成発展)する)」と定義しました。これも思考上きわめて理解しやすく、簡潔にその本質をついていると考えられます。
この定義を読むと、歴史性と偶然性がストンと腑に落ちてしまいます。つまり、歴史性(時間)がどのような意味を持つのか、偶然性が具体的にどのように展開するのか、なんとなくわかったような気分になり、それゆえの我流解釈の錯覚に陥りやすいものと思えます。
しかし現実的に生物のどのような変化(偶然性)が古生物の進化(必然性)となるのかについてはまえに書いたように推定、希望の域を出ない、というのが現実ではないでしょうか。
井尻さんもまた古生物学を主体に、地質学、解剖学、実験生物学などを経てこの考え(学説)にいたりました。
余談:井尻先生が、歯の形が顎の形と関係するという移植実験を生物学者に先駆けておこなったことは、比類なき古生物学者であることを物語っています。その後、遺伝子を扱う、あるいは分子生物学を扱う古生物学者はいるようですが、古生物からヒントを得て、当時(今も?)世界最先端の課題の実験を行った例を私は知りません。
問題点
次に上記2つの大命題=学説に従って個体発生と系統発生その問題点を、具体的にみてゆくことにします。
Haekel説の検討
まず、Haekelの考えを簡単に理解するためには彼の発生の比較の図をみるのがもっとも良いと思われるので上の図を参照してください。
この図から解ることはまず、発生の初期ほどすべての動物が似ている、動物の初期形態が類似することです。そして進化した右の動物ほど発生が進むに従って共通点が狭められています。ここから発生の過程は進化の過程の反映だというかれの意見が納得できると思います。(これについては当時から、胚の形態が似ている、つまり成体が似ているわけではないなどといいう批判もありました)
この現象は、頭尾の分化、鰓弓の分化、腎臓の分化などで系統発生が繰り返されるような現象が数多くあり、先祖型の器官が発生において一過性に現れて消失する痕跡器官であるということが納得しやすいと思います。このことから個体発生に系統発生が反映していることがあるのは間違いありません。しかし、全てかどうか、そして発生の順序となると別の問題になります。
たとえば私達の手と足、つまり上(前)肢と下(後)肢は体が頭の方向から分化するのに沿って上肢が早く分化しますが、両生類では後肢から分化することはオタマジャクシによってよく知られています。これは極端な例かもしれませんが、発生の順序は系統発生を厳密に繰り返しているとは言えない場合もあります。つまり発生の順序は種によってまた器官などによって様々だ、ということです。
では発生の現象とはどのような意味を持つのでしょう、そのためにまず発生の一般的な法則をみてゆくことにします。
だいたいどのような動物にも当てはまるのは、個体発生の初期、初期発生といわれる時期は短く、なおかつその時に体の基本的な構造が分化する、ということです。次に、それ故に大切な基本的器官は初期に形成されるため、動物に共通の器官ほど初期胚に形成される可能性が高いということです。 これに反して、初期発生ほど成体の多様性を実現するような幅広い変異が生ずる、つまり多様に変異しやすい時期でもあることです。そして、つぎに(これは重要なことなのですが)、脊椎動物では発生の順序は基本的に頭から後方へ、体幹(近位)から手足の末(遠位)へ、進むことです。私が表面的というのは、これを厳密に検討すると対称性をもって分化するためです。(これについては個体発生と系統発生の関係に重要になるため「体制の原則」としてあちこちで触れます。)
以上はあくまで脊椎動物の個体発生の法則であり、系統発生の法則ではありません。これを別の言い方に変えると、同時発生的に初期胚で様々な現象が生じるため、厳密な意味で個体発生の順序と系統発生の順序と対比させるには困難な点もまた多い、ということです。
注 参考のために発生学者のvon
Beerの発生のまとめを次に挙げております。
(http ://ja.wikipedia.org/wiki/を引用)
Von Beerの訳(進化論者ではない、Haeckelを批判)
1 大きな動物群に共通な形質は、特殊なものより先に形成される。
2 形態的に一般的なものから特殊なものが形成される。
3 一定の動物形に属する胚は、一定の諸形態を経過すると言うより、むしろそれから離れてゆく。
4 高等な動物の胚はほかの動物に似ているのではなく、その胚に似ている。
それ故に、発生初期に、基本的、根幹、根底的には系統発生を反映しているとしても、それは多くの変異の一つである、とも言えるのです。
よって、系統発生と個体発生の関係は研究者の立場(つまり古生物学、動物学、植物学、生化学、遺伝学等々)によってそれぞれ都合の良いように解釈しているようです。というよりどうしてもそのようにならざるを得ないようです。
とくに古生物学者と生物学者の捉え方は全く違っています。その原因は前にあげた時間をそれぞれの感覚で計るために系統発生と個体発生はまったく違う現象であることが本人の思惑とはべつに、理解されていない、錯覚しているということだと感じています。
現在の生物の現象が歴史的な長い時間をかけて作られた結果であるにしても、歴史的現象の短縮、あるいは現生生物の現象の繰り返しや延長が歴史的な現象となると捉えてしまうこと、これは彼があれだけの観察に自信を持っていたことでしょうが、ダーウィンに影響された錯覚だと私はみています。
もう一つ、化石の時間と現生生物の時間にかんする錯覚の例をあげれば、あとで説明を加える恐竜の絶滅の問題です。古生物学的に急激に絶滅したというのは数千万年単位の時間ですが、生きた生物を扱っていると今日明日にでも絶滅するという感覚で、せいぜいは百年や十年単位で考えてしまいます。立場によってちがう捉え方をしてしまいます。
つまり発生初期の現象が先祖的過程の反映かどうかは唯一化石によって証明されるものであり、慎重に見極める必要があるということです。
井尻説の検討
つぎに私の師匠である井尻さん(先生と呼ばれるのをとても嫌いました)の命題を検討してみます。ここで問題となるのはやはり「時間」です。
個体発生の項で説明したように、たとえ遺伝子が変わったとしても形の変化にたどり着きさらに細胞または体が安定化するためには膨大な代謝過程が必要で、その過程自体また膨大な数の変異があるのです。このような遺伝子と安定的形質との関係を確定的に説明できる研究はまだ一つもないと言って過言ではありません。それ故にES細胞、iPS 細胞等々が試され侃々諤々の議論となっているわけです。
これは発生の原理が明確となっていないことがその原因の一つとなっているのです。遺伝子なり細胞因子なりある一つの要因が判明したという研究成果は原因の一つである可能性は高いのですが、他の要因もまた沢山ある可能性を否定しえないということです。生物の生涯でその要因が超長期的に繰り返し系統発生に結びつく、あるいは反映する可能性は、まったく未知数なのです。
ですから細胞であれ個体の変化(=偶然性)が必然性(歴史的)へ至る過程、つまり現生生物の現象と化石の証拠の関連性は「推定」の域を出ない、100%の可能性がない限り、1%でも99%でも実現性という点からは同じ不可能性があるということです。
生物の変異が長い時間をかけて進化に結びつく可能性というのは、可能性としては考えられても、現実の実験では立証不可能な推定です。現在時間の生物の現象の繰り返しをそのまま万年に引き延ばせば系統発生になる、というのは推定、それも希望的推定と言わざるを得ません。
ですから井尻さんも「ちょっとずつ変わる」、と言っているのです。ちょっとずつ変わるのは化石であきらかですが、生物学的根拠はありません。長鼻類の鼻が長く進化するのは、ちょっとずつ突然変異が起きる、それも遺伝子が方向性を持って変化する、これはどうしてなのでしょう?
つまり、井尻さんの命題、論理性と歴史の一致は、哲学的な結論としては妥当であり胸にストンと落ちる結論であっても、それを担保する古生物学と現生生物学の法則はなにかという現実的な問題は解決しない、と私は考えています。
そして、「時間」の問題はわかっていると錯覚をひき起こしやすいものなのです。というより、かなりやっかいな錯覚であり、時間の感覚はなかなか掴みにくい、それゆえに私は慎重になりたいのです。
繰り返しますが、生物学者はなんとなく個体発生が何万年繰り返すと考え、また古生物学者は古生物の個体発生は現在の生物と同じだ、と考えているように私にはみえるのです。 それゆえに、進化の原因に突然変異が重要だ、とか、恐竜の絶滅の原因は星が地球へ衝突したためだ、という劇的な要因説が生まれるのではないでしょうか。
しかし、突然変異は遺伝子的には劣性遺伝子のことが多いし、星の衝突による恐竜の絶滅は地史学的には突然でも、何千、万年もかかっているのです。(現在の惑星衝突説の議論は、衝突後に地球環境が変化し、これに恐竜が適応しなかった、というようなものですが、それでも本質は変わりません。)
私はこれらの説を否定はしません、しかしいろいろな生物の系統図を振り返ると進化は少しずつ進むし、恐竜の絶滅には星が衝突して環境が変化したにしても、何百、千万年もかかるのは生物側(主体)にも原因がある、と考えるのが自然ではないでしょうか。
少しずつ進化するには少しずつ変異が生じ、この変異が地史学的時間つづかなければなりません。
この問題に関する私の考えは、「はじめに」で書きましたが、個体発生と系統発生の関係を担保するものは「体制の原則」であるという結論です。体制の原則に沿った変化は長長期的に続く可能性がある、というものです。
系統発生と個体発生の関係をどのように捉えるか
では、系統発生(進化)と個体発生の関係をどのように捉えたらよいのでしょうか。その違いをつぎに挙げてみます。
まず第一点は、系統発生と個体発生は全く違うものであるということを強く意識したいと思います。その根拠は時間のスケールが違う、という当然のことです。
第二点は、系統発生は個体発生に反映していると言うことです。
この根拠は化石で証明されている裏付けがある発生途上に現れて消失する痕跡器官に現れている、と考えられます。例えば、鰓弓、腎臓の分化などなどです。同時にこれは個体の変異の一つででもあるのです。
第三点は、個体発生の順序は、必ずしも系統発生の順序を反映しない、ということです。体幹形成から頭部が分化し、四肢が分化すると言う順序、あるいは、腎臓の分化などなどはあるていど系統発生の順序を反映していると推定されますが、発生初期は非常に早く幾多の分化する時期でもあり変異もまた数多くおこるため必ずしも系統発生の順序と断定できなないのです。
繰り返しますが、それゆえに系統発生と個体発生の関係を取り扱う研究には十分根拠を示し、慎重な態度で臨むべきだと私は考えています。
系統発生と個体発生のおおきな相違点をまとめると次のようになります。
1 系統発生では発生、変化、繁栄、滅亡の四段階、個体発生では発生、変化、成熟、死の四段階とおおまかに区分できます。
2 このうち発生と滅亡は化石では明確でないことがあり個体発生とは違います。
3 系統発生の変化と繁栄過程、個体発生の変異と成熟過程は、形態と数の点で共通の型といえます。ただしそれは特定の種類であり全てではありません。
4 系統発生と個体発生では時間のスケールが全く違います。
5 系統発生、進化の結果が現在の個体(個体発生)です。
6 系統発生と個体発生とが類似する現象、つまり反映している現象は沢山あります。
系統発生と個体発生が類似しているということは事実です。だから相互に反映していることも確かだ、と考えられます。そして古生物学的事実はゾウの鼻が長くなった例を上げるまでもなく、少しずつ変わって進化した、のです。
そのとおりなのですが、私はこれだけでは満足できません。
何故、少しずつの方向性のある変化が何千、何万、何十万、何百万年、いやそれ以上続いて繰り返された要因、そこには幾多の変異があり生き残ったのは僅かだとしても、生き残る根拠、要因は明らかになっていません。もっと厳密に言えば生物自体の要因が明らかになっていない、ということです。だから連続変異説や、「相同性」や「相似性」等々の議論が尽くされてきたのです。
ここで考えなければならないことは、古生物学的事実と環境の事実、これを無理なく整合的に捉えることが必要である、ということです。
詳しくは「7 変異」についてと「8獲得形質」さらに「12 進化の定義と私の進化理論」で検討を加えましたが、ここでは要点のみ紹介いたします。
結論から言えば、変異というのは遺伝子のみではなく、DNAの複製や転写、また蛋白などの産生、分泌、これらに関わる酵素、補助酵素など全てにみられる現象で、生物学的な全過程、全階層に起こっていることが分かっています。ですからゾウ(長鼻類)の鼻が長くなる方向への進化は、鼻が長くなる変異が超長期にわたって少しずつ続いてきた結果です。長鼻類全体に鼻が長くなることからいえるように、この種類特有の現象です。つまり、他の動物は同じ環境でも他の方向へ進化したわけです。
同じ環境でも異なる進化の原因は、種の特異性といっていますが、これを引き起こすのは「嗜好性」です。どのような生物でも全階層に嗜好性を持ちます。それ故に多様性が生まれ、種全体の集団的嗜好性が種の特異性となり、種が形成されるわけです。これは自然界でも同じであり、ある物質特有の性質などと表現されます。ですからこれは普遍的性質であり「体制の原則」の一つだといえるのです。
嗜好性を科学的ではないという批判を沢山受けました。これは同じ環境でも同種類で違う変異のおこる原因ですし、類似した体質を持つ集団(種など)では共通の嗜好性があるのです。類は友を呼ぶ、のです。
生物の進化を振り返り理解できることは、第一に単細胞から多細胞へ、複雑な構造への道(階層の増加)、第二に原始の海から陸へと拡大したことが読み取れます。そして、現在の生物をみると、生活圏は数千メートルの海の底から数千メートルの山の頂、否、ある種の花粉は1万メートル以上も飛ぶと言われています。またある種の菌は地中深くに生息し、また耐熱菌などはかなり高温に生息することもわかっています。これほど多様な生活圏をもつのはそれぞれの嗜好性による生物(生命)の進化の結果と捉えてよいと考えられます。
残念ながらこのような要因の議論はいままでない、というか私は知りません。
つまり、長鼻類全体の連続性を持った進化を保証する、担保するのはこの体制の原則だと、私は考えています。
この様なことから系統発生と個体発生の関係はつぎのように要約することができます。
個体発生(これ以外の代謝も含めて)の変異のうち「体制の原則」に沿う変異ほど体内外と調和して完成する(表現型となる)可能性が高く、超長期に繰り返す可能性があり、系統発生に反映する。またそれ故に系統発生は個体発生に反映している。
井尻さんの命題である「個体発生は系統発生によってあたえられ、系統発生は個体発生を通じて展開する。」つまり、「個体発生の合法則性(論理性)は、系統発生の必然性(歴史性)によってあたえられ(獲得され、刷り込まれ)、系統発生の合法則性(論理性(=必然性)は、個体発生の偶然性(変異性)を通じて展開(生成発展)する)」の「必然性(論理性)」「偶然性(変異性)」は、「体制の原則」という共通の基盤によって理解できると考えられます。
この多様な進化をなしえるようにする(担保する)のは、体制の原則に沿った変化(=進化)、つまり全地球的環境とその変化と親和性があるから、というのが根拠です。